For several decades, the exciting question has been at the centre of scientific research on whether and how aging processes can be influenced. In the following therefore, the question should be investigated to what extent the increase in the ATP level (ATP = adenosine triphosphate, the energy molecule of life) can influence the life extension of a biological organism - here a plant. Furthermore, it should be investigated whether scalar waves, which have been de-scribed as carriers of energy and information in many publications [1], are able to cause an ATP increase in the scalar wave plant.
Scalar waves are two of three possible wave types, the electromagnetic wave, in 1888 discovered by Heinrich Hertz, the electric scalar wave, discovered by Tesla around 1890, and the magnetic SW, which plays the crucial role in biology [2]. However, until now the scalar waves have not been recognized by the physicists because they are not describable with the Maxwell equations. It is important to know that the third equation (div B = 0) was extended by the Helmholz Society in 2009 (div B> 0) by the discovery of Magnetic Monopoles. Unknowingly, this discovery results in scalar waves.
While the Hertzian waves have a transversal-wave characteristic, whose propagation front separates spherically from the sender with fundamentally quadrat-ic decreasing of energy density, the scalar waves are longitudinal waves, which are based on a direct trajectory from the sender to the receiver, the energy emitted by the transmitter is completely collected again by the receiver. This applies, however, only in the so-called resonance between transmitter and re-ceiver, which requires the same waveform, the same frequency and the opposite phase, but which was technically already considered as a rule. This is also a necessary condition for the energy transfer, which Tesla had already demon-started and which cannot be represented with the Hertzian waves because of the quadratic decrease.
In Figure 1 the characteristics of the wave types are compared.
Properties of the wave types |
Transverse wave |
longitudinal wave |
Electromagnetic Wave (Hertz) e.g. Radio wave |
electrical or magnetic scalar wave e.g. Radio wave energy (Tesla) or DNA (Meyl) |
Energy dissipation |
Energy conservation |
Propagation Spherical with v = c (= constant) |
towards the electric or magnetic Field pointer with v (= arbitrary) |
Figure 1. Comparison of the properties of both types of waves
The so-called magnetic wave first discovered by Meyl, in which the directional vector of the wave coincides with the magnetic pointer, is typically used in the biological context [3-5], while technical applications mostly use the electric wave already used by Tesla (Tesla wave).
Nature uses the magnetic scalar wave instead of the electrical Tesla-wave. This can also explain why it is possible for up to 10000 different biochemical transactions to occur in each cell every single second, a process that cannot be explained by the conventional transaction model of mainstream chemistry.
As shown in the book "DNA and Cellular Radio" [2], the DNA is an important transmission-way of scalar waves, as also are the so-called microtubulies in the plant cells. Thus, one can assume that the cell not only communicates intracellularly with the help of its own generated scalar waves, but also intercellularly with other cells throughout the whole organism.
As a scalar wave (SW) source for the experiment a more recent development of Meyl's experimental device, the so-called 'cell radio', was used at a power of 7 mW for 90 seconds. See (Figure 2) left picture.

Figure 2. Parts of the experimental setup with the 'cell radio' (left), handle with flowers (top right) and ATP Lumitester PD-30 from Kikkoman (bottom right)
The plant buds were supplied with scalar waves using a single pulse (90sec) at about 6.7 MHz. As parameters for the aging process, the diameter of blossom (in cm) and the ATP content at the different stages of the flourishing/aging process (Figure 3) were measured in relation to their weight (in g, without stem and sepals).

Figure 3. Four typical flowering stages of Ipomea purpurea
Each blossom was individually weighed and macerized in 300ml of distilled water using a blender (18000 rpm, 2 min). Subsequently, the ATP content was determined for an aliquot of 100µl, according to the luciferine / luciferase system (Flexikon.doccheck.com/de/Luciferin-Luciferase).
Results
Different flowering stages of Ipomea purpurea and their ATP content
Ipomea purpurea is a winding plant with impressive trumpet-shaped blossoms in different colours from red to purple and magenta, flowering between July and September (Figure 3) shows that flowers of different stages (phases 1 to 4) are present at the same time on one bush. Since the flourishing process from the opening of the buds to the wilting stage lasts only one day, it offers an impressive model of the aging process.
Initially, the flowers were harvested in the field at the bud stage before the 24-hour flowering process was monitored under water supply and room conditions. Still in the bud stage they were subjected to the corresponding SW treatment mentioned above, for 90 seconds in contrast to controls, afterwards they were processed according to 4 different stages cf., (Figure 3). The flowers at bud stage (phase 1), at full blossom (phase 2), at the wilting stage (phase 3) and at totally wilted stage (phase 4) were weighed after separation from stem and sepals and - as already described - processed for ATP measurement.
Figure 4 shows the decrease of ATP during the different flowering stages, from 111,000 RLU/g at the bud stage (relative light units, corresponding to approximately 2µmol ATP/g) to 55,000 RLU/g at the wilted stage (about -50%). This result is explained as an energy consumption process necessary for blossom opening (phases 1 and 2) determined as an ATP-decrease from phase 1 to 2. During aging (wilting) the ATP-level is reduced again (phases 2, 3 and 4) characterizing the aging process itself.
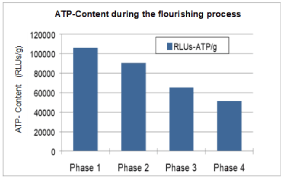
Figure 4. During the process of flowering and wilting the ATP content is decreased by 50% compared to that of the bud (phase 1, phase 2 = full flowering, phase 3 = wilting, phase 4 = wilted stage; RLU = relative light units)
Different flowering/aging stages of Ipomea and corresponding ATP levels after SW treatment
In the next step controls (without SW treatment) were compared with SW-treated flowers (treated once with a single SW pulse 90 sec, 7 mW). The experiment was started at 5pm using buds (Figure 5).
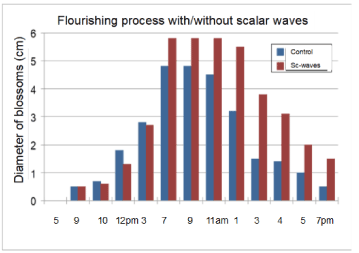
Figure 5. The 24-hour flowering/aging process of the Ipomea without (blue) / with (red) scalar wave treatment. The aging process of the control was started at 11am, that of SW-treated one at 1pm, i.e. 2 hours later
The diameters of the blossoms were determined using the largest axis (in cm), both of them were almost equal at 12pm. However, at 3 am, the diameter of the SW-treated plants was increased up to 20% in contrast to the untreated ones.
Figure 6 indicates the difference of the wilting process of treated and untreated plants SW-treated plants start to wilt at 1pm (Figure 6) top, middle, whereas controls begin at 11am (Figure 6) top left, that means a difference of 2 hours. Moreover, the different blossom diameters of 20% are obvious (Figure 6) top and lower row. Figure 5 and Figure 6 show that the extension of the flourishing process was about 10% after SW-treatment.

Figure 6a. Flowering stages of Ipomea in dependence of a 24 hours rhythm.
Left side glasses: Ipomea without SW treatment (notice the red colour), right side glasses: SW-treated plant (violet colour)

Figure 6b. Flowering stages of Ipomea in dependence of a 24 hours rhythm.
Left side glasses: Ipomea without SW treatment (notice the red colour), right side glasses: SW-treated plant (violet colour)

Figure 6c. Flowering stages of Ipomea in dependence of a 24 hours rhythm.
Left side glasses: Ipomea without SW treatment (notice the red colour), right side glasses: SW-treated plant (violet colour)
Another symptom during the aging process was the colour change from red to deep violet in SW-treated Ipomea (Figure 6) top, right glasses. However, finally this colour difference is compensated during the wilting process when comparing right glasses with left ones (Figure 6) lower row.
Both Ipomea plants (treated/untreated), harvested from the same shrub, showed the same colour (red) at the beginning of the experiment, but the SW-treated plants not only changed their colour to deep violet, but also increased the size of the blossom at the same time by 20% compared to the control. In addition, the wilting process after a single SW pulse was delayed by 2 hours, corresponding to an extended overall lifespan of the blossom of about 10%. The different colours and their interpretation will be discussed later.
Additionally, the ATP-levels were measured at each of the 4 stages of the flourishing process with/without SW-treatment. After macerisation of the blossom the ATP levels were determined in an aliquot of 100µl.
Figure 7 shows that the SW treatment of the bud increases the ATP-level by 40% , which however decreases afterwards to a final ATP-content at the wilted stage, which is identical then with that of untreated plants. This initial energy difference of 40% is consistently ‘consumed’ along the opening movement of the SW treated buds as well as the extended wilting process compared to the untreated flowers, resulting in an extension of the lifespan by about 2 hours. Thus, the initial excess-energy is used up for an extended flowering process and a lifespan extension by about 10%.
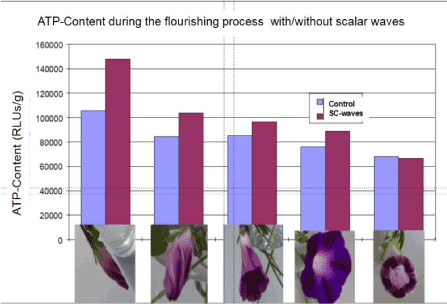
Figure 7. The ATP-level in the buds is increased by 40% using SW-treatment (90sec; 7mW). This ATP difference is metabolized during the flowering process thus delivering energy for a prolongation of the flourishing process by 2h (almost 10% of the life span)
Since ATP production takes place in the mitochondria, it can be assumed that the mitochondria are the sites of action for the scalar waves. Thus, a hypothesis about the mechanism of action should begin here. Mitochondrial ATP formation is common to all living creatures, whether plant, animal or human, since evolution has (probably) taken this step, i.e. inclusion of archaea bacteria living in the oxygen atmosphere.
The essential biochemical process of ATP production in the mitochondria is the respiratory chain, where reduction equivalents NADH [6] via co-factors reduce the oxygen from the air to water (Figure 8).
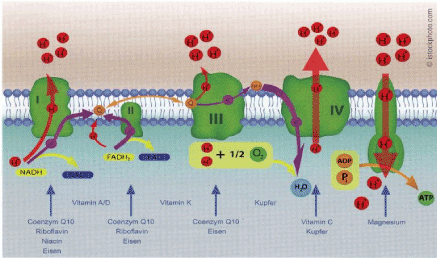
Figure 8. Respiratory chain in the inner mitochondrial membrane (Source: Lemperle, M., Kesten, S., OM&Ernährung, 2017, 159, p.23)
To synthesize ATP, four defined enzyme complexes transport electrons through the inner mitochondrial membrane, which at the end of the chain reduces oxygen to water. The simultaneously released protons are pumped into the intermembrane space. The resulting proton gradient is the prerequisite for the formation of ATP via the ATP synthase complex.
The presence of a number of co-factors is necessary for the functioning of the mitochondrial respiratory chain: in complexes 1 to 3, e.g. CoQ10 (ubiquinol) plays a key role as an electron donor (Figure 8). Without ubiquinol, energy in the form of ATP molecules is not sufficiently provided to the body cells. The undesirable consequences are diseases and premature aging [7,8]
If this respiratory chain does not work sufficiently at the 4 complexes, resp. a single mechanism is blocked as a result of diseases, aging processes or drugs, environmental toxins, electro smog, nitrosative stress [9,10], the reduction to water may be interrupted and intermediates like the reactive oxygen radicals (ROS) are built up as the source of oxidative stress.
The SW-induced colour change of the Ipomea
A last point should be mentioned in this context, which focuses on the colour change of the Ipomea after SW-treatment. This change from red to purple (Figure 9) left, top is due to the presence of anthocyanins in the Ipomea, plant polyphenols responsible for the strong colour of the flowers.
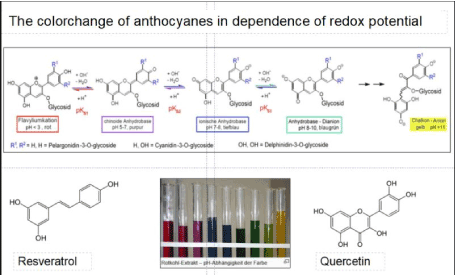
Figure 9. Redox state of anthocyanins: colour changes as a function of the phenolic or the quinoid state of the anthocyanins (see top row, left: red (phenolic state) purple (quinoidal state).) In the test tubes below, the colour change is shown (depending on pH) (Source: https://en.wikipedia.org/wiki/Anthocyanin)
The scalar waves are hypothesized to release electrons from polyphenols, oxidizing them with the result of a colour change. The transition from the phenolic red coloured, reduced state to the quinoid purple, oxidized state is obvious. In the course of the wilting process, the purple/ violet state changes back again to the red one (Figure 6). On this basis, it is assumed that SW-treatment enhances electron flow in the respiratory chain of the mitochondria thus increasing ATP level in the tissue of the plant (Figure 7).
The SW pulse is hypothesized to be absorbed via mesomery, typical for organic ring molecules of plant polyphenols, such as the anthocyanins. Polyphenols - like resveratrol and quercetin (Figure 9) are assumed to function as 'antenna molecules' absorbing, storing and releasing the energy of SW pulses. Subsequently, these energy pulses seem to be transferred via co-factors, such as CoQ10, within the respiratory chain thus stimulating the electron flow. This phenomenon is correlated with an increased ATP level, which optimizes the functionality of the mitochondria and thus may extend the lifespan of organisms.
Conclusion
In summary, SW-absorbing plant polyphenols (i.e.anthocyanins) seem to activate oxygen-dependent mitochondrial activity increasing ATP-levels. This, as the experiments have shown, can have a lifespan-prolonging effect on plants. Observations in the past have already shown that polyphenols have lifespan-prolonging effects via the so-called SIRT-mechanisms [11-14].
Further experiments will certainly contribute to clarify this complex of questions in the future.
- University of Bonn, IMBIO (Institute of Molecular Physiology and Biotechnology of Plants, Bonn), Karlrobert-Kreiten-Str. 13, D-53115 Bonn.
2021 Copyright OAT. All rights reserv
- University Furtwangen, First Tranfercenter for Scalar wave technology, Erikaweg 32, D-78048 Villingen-Schwenningen.
References
- Meyl K (2012) Cellular Communication, signaling and control from the different perspectives of Biology, Chemistry and Field Physics, WMSCI 2012 Proceedings Vol.II, page 113 – 117, Chair of BMIC/ WMSCI 16 th World Conference Orlando, Florida, USA, July 18, 2012. http://www.iiis.org/CDs2012/CD2012SCI/SCI_2012/PapersPdf/BA861JU.pdf
- Meyl K (2010) DNA und Zellfunk“, Indel Verlagsabteilung.
- Meyl K, Schnabl H (2015) Biological signals transmitted by longitudinal waves influencing the growth of plants, SEEK Digital Library, Int. Journal of Environmental Engineering - IJEE 2: 23-27.
- Ebbers JA, Meyl K (2014) Drug effects in yeast mediated by scalar waves. Medical Science 8: 58-62.
- Meyl K, Schnabl H (2015) Biological signals transmitted by a resonant L-C-oscillator, PIERS draft proceedings, progress in electromagnetic research, prague, Czech Republic, 2601-2606.
- NADH: Birkmayer, G.D. 2017, ISBN 978-3-200-04066-3.
- Kuklinski, B.,Mitochondrien, Aurum Verlag, 2017, S. 30-38.
- Schmidt N, Schmidt E (2018) OM&Ernahrung 2018, Sonderheft Mitochondrienmedizin, S.159-63.
- Lemperle,M und Kesten S (2018) OM&Ernahrung 159: 20-259.
- Hüther H (2018) OM&Ernahrung, 2018, Sonderheft Mitochondrienmedizin,S. 57-59.
- Wood (2004) Nature 430: 686-689.
- Porcu M, Chiarugi A (2005) Sciences 26: 2-12.
- Sinclair DA, und Guarente L (2006) Spektrum der Wissenschaft.
- Kremer H (2007) OM und Ernahrung 119: 1-5.