Two hundred years ago (early 1819), the French naturalist Jean-Baptiste de Lamarck (1744–1829) lost his eyesight and had to cope with poverty over the last decade of his life. In the previous year (1818), Lamarck had introduced the term “Hirudinea”, and described all the leech species known at that time in one of his books. Here, we recount the life and achievements of Lamarck with reference to leeches of the genus Hirudo. We document the evolutionary origin (somewhere in Asia, ca. 15 to 20 million years ago), occurrence, speciation patterns, systematics and practical application of these parasitic annelids. It is concluded that Lamarck’s pioneering work on the systematics of invertebrates provided a solid basis for a research program into the evolutionary biology and physiology of these important model organisms.
Lamarck, leeches, Hirudo-therapy, Hirudo medicinalis, evolution
Carl Linnaeus (1707–1778) was a world-class naturalist who gave living organisms two Latin names, and described hundreds of new species of plants and animals. Among them, the earthworm (Lumbricus terrestris L. 1758) and the medicinal leech (Hirudo medicinalis L. 1758) are well-known invertebrates that are members of the phylum Annelida. Both earthworms and leeches are protandric hermaphrodites that act, during sexual reproduction, first as male (distributor or sperm), and then as female (provision of egg cells, followed by cocoon production). The 18th century Linnaean system of the “classis Vermes”, with the three dissimilar orders “Intestina, Mollusca and Testacea”, was later replaced by the much more sophisticated concept of Jean- de Baptiste Lamarck (1744-1829).
This eminent French naturalist published numerous papers and monographs on the classification of the “lower animals”. Moreover, Lamarck was one of the first scientists to introduce the term “biology” and proposed the idea of organismic evolution [1]. In 1818, Lamarck coined the word “Hirudinea”, and described-classified all leech species known at that time (Figure 1).

Figure 1. Reproduction of the title page of Lamarck’s Vol. 5 in the series “Histoire Naturelle des Animaux Sans Vertèbrates”, published in July 1818, supplemented by a portrait of the author. In this book, the term “Hirudinea” (leeches) was coined.
In this article, we describe Lamarck’s pioneering work in this area of invertebrate zoology. In the second part of our account, we summarize the current status of the systematics of medicinal leeches (members of the genus Hirudo), invertebrates that are both of theoretical and practical value [2]. Finally, we discuss the question as to whether or not, two decades after Lamarck’s description of H. medicinalis, this species still exists in European freshwater ecosystems.
Lamarck’s achievements and legacy
Two hundred years ago (March 1819), Lamarck’s fame rapidly declined, which is, at least in part, attributable to his controversy with the creationist Georges Cuvier (1769–1823). This Biblical literalist (and gifted comparative anatomist) attacked Lamarck’s ideas on the gradual transformation of animals to such an extent that the reputation of the “atheistic evolutionist” was considerably damaged. In addition to this professional disaster, the hard-working biologist lost his eyesight. As described in corresponding monographs on the life and scientific work of this genius, Lamarck had to cope with poverty over the last decade of his life and, for reasons not yet known in detail, became blind [1]. In this section, we briefly summarize his lasting impact on modern biology.
The French biologist Lamarck was the “true father” of a concept we today call “naturalistic evolution”. As detailed in a classical biography [1], Lamarck was the first scientist to disregard the then-popular idea of “Independent Creations” of all forms of Life, as described in Genesis of the Bible. As an alternative, Lamarck published, in 1809 [3], his famous theoretical concept of the “transformation of species”, with reference to a few speculative “primitive” unicellular organisms that may have existed a long time ago.
Despite these insights, Lamarck is today not so much remembered for his “discovery of evolution”, but because he suggested a kind of “inheritance of acquired characteristics”, as detailed in his Zoological Philosophy [3]. However, Lamarck was also a creative botanist and invertebrate zoologist. Before he took over a position as Professor of “lower animals” in the Natural History Museum of Paris (France), these neglected organisms were largely unknown to science. Therefore, it is fair to say that Lamarck was the founder of invertebrate zoology [4,5].
Two decades ago, in Volume 5 of a series of books on the systematics of invertebrates, Lamarck (1818) introduced the class “Hirudinea” (leeches) (Figure 1). In this monograph [6], he listed two European species, Hirudo medicinalis, and “H. sanguisorba”. The type species of the class Hirudinea Lamarck 1818 (sangsue médicinale, i.e., the medicinal leech), is still accepted today and regarded as a valid species (Figure 2). However, Lamarck’s second taxon, “H. sanguisorba”, remains an enigma. No credible leech biologist has, to the best of our knowledge, provided any evidence so far as to the systematic status of this Lamarckian leech taxon. In addition to these two Hirudo-species, Lamarck (1818) listed a number of other taxa as members of his newly established class “Hirudinea”.
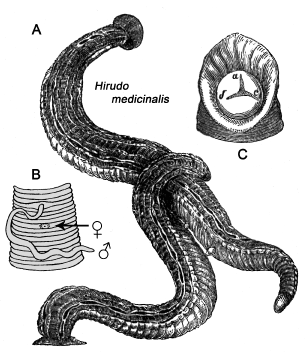
Figure 2. Pair of medicinal leeches (Hirudo medicinalis) in copulation (A) and schematic drawing of the male copulatory organ (everted penis), separated by three annuli by the female gonopore (vagina, i.e. copulatory organ) (B). The drawing is supplemented by a schematic rendering of the mouth, showing the three teeth of these hermaphrodites (C) (adapted from anonymous drawings, ca. 1890).
A “polymorphic species” consists of two distinct taxa
As a result of the use of leeches in phlebotomy (bloodletting) throughout Europe (a peak was reached ca. 1850), numerous “varieties” of the European medicinal leech “H. medicinalis” were distinguished by practitioners and biologists alike. In a classic monograph on leeches, Mann [7] adopted this interpretation and wrote that H. medicinalis must be regarded as a “highly variable species”. Accordingly, Sawyer, in his influential three-volume-book [8], wrote that all colour (and pattern)-variants of European medicinal leeches should be assigned to the polymorphic taxon H. medicinalis Linnaeus 1758 (syn. H. officinalis Savigny 1822). However, based on detailed analyses of geographical distributions of leech populations throughout Europe, pigment patterns, and DNA-sequences, it was shown that H. officinalis is not a “colour variant of Linnaeus’ type species”. Rather, it represents a separate taxon, the Mediterranean medicinal leech H. verbana Carena 1820 [9-15]. Both species reproduce by reciprocal insemination via copulation of two fertile hermaphrodites, and deposit their cocoons into moist soil (Figure 2). From these egg sacs, which are characterized by a complex fine structure that protects the cocoons from desiccation [16], juvenile leeches hatch that show the species-specific colour pattern of their parents.
Hirudo medicinalis (Figure 2), and H. verbana (Figure 3) are used in Europe for bloodletting and in biomedical research programs. However, as detailed elsewhere [11,15], today we know that “H. medicinalis” represents a group of closely related, reproductively isolated biospecies. This important insight is summarized in the next section.
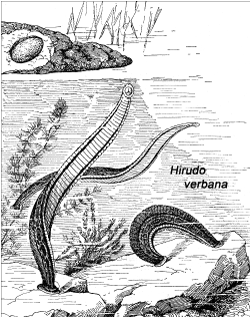
Figure 3. The Mediterranean medicinal leech (Hirudo verbana Carena 1820) in its natural habitat. Note that the cocoons are deposited in moist soil on land (adapted from an anonymous drawing, ca. 1920).

Figure 4. The European medicinal leech (H. medicinalis), represented by a relict population in Germany. The stagnant aquatic ecosystem contains numerous plants and amphibians. The inset shows a group of adult H. medicinalis that are alerted by water movements caused by a person who investigated this population of leeches (adapted from ref. [20]).
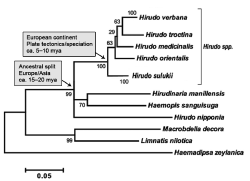
Figure 5. Maximum Likelihood phylogenetic tree, based on a combined COI, 12S and 18S-mt-DNA-data set (1,514 total positions). European Hirudo species form a distinct clade with H. sulukii as a basal member. Bootstrap values are indicated. All six species of the genus Hirudo are shown, with Haemadipsa zeylanica, Limnatis nilotica and Macrobdella decora as outgroups. In addition, the biogeographical processes that have led to the separation of Asian and European leech species are indicated (adapted from ref. [17]).
Hirudo medicinalis: An evolving species complex
During the 19th century, most zoologists regarded Hirudo medicinalis L. 1758 as a “polymorphic leech taxon” [7,8]. However, as described in a recent monograph [17], it has long been suspected that “H. med.” consists of a Northern and Southern population that may represent separate species within the genus (i.e., H. medicinalis L. 1758 and H. verbana Carena 1820). Extensive biogeographical studies and the collection of hundreds of representative specimens from different localities throughout the Palearctic region (Figure 4) yielded surprising results. Detailed molecular analyses of these collected specimens, based on DNA-sequence information of combined cytochrome oxidase-sub unit 1 (COI) and 12S/18S-data sets, revealed that the genus Hirudo consists of at least 6 reproductively isolated (true) biospecies [17,18].
In addition to the above-mentioned “classical” taxa (Hirudo medicinalis L. 1758 and H. verbana Carena 1820), which are listed under the names “European vs. Mediterranean medicinal leech”, respectively, and the North-African “Trout (or Dragon)” leech (H. troctina Johnson 1816), more recently three further species were discovered (Figure 5): The Korean blood-sucking leech (H. nipponia Whitman 1886), the “Persian (or Georgian)” medicinal leech (H. orientalis Utevsky and Trontelj 2005), and the Turkish medicinal leech (H. sulukii Saglam, Saunders, Lang and Shain 2016).
As the phylogenetic tree depicted in Figure 5 shows, we have to distinguish between the closely related species H. verbana, H. troctina, H. medicinalis, H. orientalis and H. sulukii, a clade that originated ca. 5 million years ago, and the more distantly related Korean species H. nipponia. Interestingly, H. nipponia is a sister taxon of the well-known species Hirudinaria manillensis (Asian medicinal leech) and H. sanguisuga, the so-called “horse-leech”, a taxon displaying a world-wide distribution [19]. We suggest that H. sanguisuga may have been misclassified, but more work is required to corroborate this hypothesis [17].
The genus Hirudo appears to have originated somewhere in Asia during the Lower Miocene (ca. 15 to 20 million years ago, mya) and thereafter dispersed eastward (i.e., toward Japan; H. nipponia) and westward towards Europe. The Euroasian Hirudo lineage speciated between 5–10 mya as a consequence of several geological events (e.g., the Zanclean flood, formation of Levantine land bridges, building of the Taurus Mountain chain), which effectively subdivided Europe into geographic regions that restricted hybridization (i.e., gene flow) between populations [17]. The closely related, extant species of Hirudo – H. medicinalis, H. verbana, H. troctina, H. orientalis and H. sulukii, – are currently observed naturally across the Eurasian landscape, with some mixing as a consequence of anthropogenic activity (e.g., farming) [11,15].
Occurrence of H. medicinalis in Europe
A key question in biodiversity research is whether or not the type-species H. medicinalis still exists as viable populations throughout the Northern parts of Europe. Fortunately, a number of recent reports have shown that this famous annelid, referred to and described by Lamarck in 1818 (Figure 1), has survived in its descendants.
In Germany, a number of large, wild populations of H. medicinalis still occur, notably in the Eastern (less populated) part of the country (for instance, the Federal State of Sachsen-Anhalt) [18,20] (Figure 4). In contrast to the “Southern” species H. verbana, which prefers aquatic habitats in steppe landscapes, H. medicinalis-populations have more frequently been discovered in ponds close to (or within) deciduous arboreal zones (for instance, Birch forests). Both species suck the blood from mammals and, notably, amphibians that are co-inhabitants of their respective aquatic biotope (Rana arvalis, Pelophylax lessonae, Triturus cristatus, etc.). In 2018, H. medicinalis-populations were discovered in Germany (Figure 4), Denmark, Norway, Sweden, Belgium, the Netherlands, France, Luxembourg, Poland, Estonia, Belarus, Lithuania, Latvia, and the Russian Federation (Southern Ural Mountains). In addition, small populations have been found in the United Kingdom, Switzerland, Slovakia, Austria, the Czech Republic, Hungary, Ukraine, Slovenia, Croatia and Romania [18,20,21]. In Turkey’s wetlands, H. medicinalis has been confused with H. verbana [12]. Hence, we do not exactly know how many relict populations of this species occur today in this country. Finally, it should be mentioned that H. medicinalis is classified as “NT category” in the IUCN Red List (i.e., “near threatened”) [18].
Unfortunately, its European sister taxon, H. verbana, has not yet been granted such a protection status. Hence, it is necessary to label H. verbana as a “sub-species” of H. medicinalis in order to provide protection for both European species. Major reasons for the decline of these Hirudo-species are over-collection for medicinal purposes, and the systematic destruction of wet land habitats, which usually leads to the loss of amphibian populations, on which the blood-sucking leeches depend [14,15].
This article was written to commemorate the publication of Lamarck’s 1818-monograph wherein the term “Hirudinea” was coined, and to remember that this world-famous biologist had to cope with private hardship over the last decade of his life (blindness and poverty from March 1819 until his death on Dec. 18, 1829) [1,4,5]. Lamarck’s pioneering work led to an historical sequence of events that have kept species of Hirudo in the public’s eye for the past few centuries [2]. These include the popular practice of bloodletting throughout the 19th century, the classic neurobiological preparation at the turn of the century, and more recently (over the past ca. 40 years), the large-scale harvesting of Hirudo species on Eurasian leech farms. These “domesticated” leeches, which are considered in Germany as “medicine”, are distributed worldwide, primarily as healing agents in reconstructive surgeries, but also as pharmaceutical targets (e.g., anti-coagulants/analgesics isolated from leech salivary glands) [14,19,22].
- Packard AS (1901) Lamarck, the Founder of Evolution: His Life and Work. Green & Co., New York.
- Shain DH (ed.) (2009) Annelids in Modern Biology. John Wiley & Sons, New York.
- Lamarck J-B de (1809) Philosophie Zoologiqué. Verdiere, Paris.
- Gadjev I (2015) Nature and nurture: Lamarck’s legacy. Biol J Linn Soc 114: 242-247.
- Weiss A (2015) Lamarckian illusions. Trends Ecol Evol 30: 566-568. [Crossref]
- Lamarck J-B de (1818) Histoire Naturelle des Animaux sans Vertèbres. Tome Cinquieme. Verdiere, Paris.
- Mann KH (1962) Leeches (Hirudinea). Their Structure, Physiology, Ecology and Embryology. Pergamon Press, London.
- Sawyer RT (1986) Leech Biology and Behaviour. Vol. 1-3. Oxford University Press, Oxford.
- Nesemann H, Neubert E (1999) Annelida, Clitellata: Branchiobdellidae, Acanthobdellea, Hirudinea. In: Süsswasserfauna von Mitteleuropa 6/2 (eds J. Schwoerbel & P.Zwick). Spektrum, Heidelberg.
- Kutschera U (2007) Leeches underline the need for Linnaean taxonomy. Nature 447: 775. [Crossref]
- Kutschera U (2012) The Hirudo medicinalis species complex. Naturwissenschaften 99: 433-434. [Crossref]
- Trontelj P, Sotler M, Verovnik R (2004) Genetic differentiation between two species of the medicinal leech, Hirudo medicinalis and the neglected H. verbana, based on random-amplified polymorphic DNA. Parasitology Research 94: 118-124. [Crossref]
- Phillips AJ, Siddall ME (2009) Polyparaphyly of Hirudinidae: many lineages of medicinal leeches. BMC Evolutionary Biology 9: 246-257. [Crossref]
- Elliott JM, Kutschera U (2011) Medicinal leeches: Historical use, ecology, genetics and conservation. Freshwater Reviews 4: 21-41.
- Kutschera U, Elliott JM (2014) The European medicinal leech Hirudo medicinalis L.: Morphology and occurrence of an endangered species. Zoosyst Evol 91: 271-280.
- Saidel WM, Saglam N, Salas de la Cruz D, Saunders R, Shain DH (2017) Elaborate ultrastructure of the Hirudo (Annelida: Hirudinae) cocoon surface. J Morphol 279: 545-553. [Crossref]
- Saglam N, Saunders R, Lang SA, Shain DH (2016) A new species of Hirudo (Annelida: Hirudinidae): historical biogeography of Eurasian medicinal leeches. BMC Zoology 1/5: 1-12.
- Surugiu V (2018) On the presence of the European Medicinal Leech Hirudo medicinalis Linnaeus, 1758 (Annelida: Hirudinea) in Romania. Travaux du Muséum National d’Historie Naturelle “Grigore Antipa” 61: 7-11.
- Elliott JM, Dobson M (2015) Freshwater leeches of Britain and Ireland. Keys to the Hirudinea and a Review of their Ecology. Freshwater Biological Association Scientific Publication No. 69, Cumbria, UK.
- Arndt E (2014) Ergebnisse der Ersterfassung und -bewertung. Ringelwürmer (Annelida). Hirudo medicinalis Linnaeus, 1758 - Medizinischer Blutegel. Berichte des Landesamtes für Umweltschutz Sachsen-Anhalt, Halle. Sonderheft 3/2014: 23-30.
- Siddall ME, Trontelj P, Utevsky SY, Nkamany M, Macdonald III KS (2007) Diverse molecular data demonstrate that commercially available medicinal leeches are not Hirudo medicinalis. Proc Roy Soc B: Biol Sci 274: 1481-1487. [Crossref]
- Trontelj P, Utevsky SY (2005) Celebrity with a neglected taxonomy: molecular systematics of the medicinal leech (genus Hirudo). Mol Phylogenet Evol 34: 616-624. [Crossref]