Abstract
Substances from terrestrial plants and marine organisms have since long been recognized as important sources of bioactive substances. This has led to the development of a large variety of drugs to treat human diseases such as, among others, a number of antihypertensive, hypoglycemic, cardiovascular, antibiotic, and antineoplastic agents. More recently, the amazing biodiversity represented by the myriad of insect species has been realized to produce an equally exceptional source of bioactive chemicals with therapeutic potential. Many of these compounds serve as highly effective defensive and predatory chemicals and have enabled insects to survive for hundreds of millions of years and to diversify to the countless different species known today. As some of these chemicals possess meaningful pharmacological properties, they represent interesting candidates for new drug discovery and development programs. A few examples are substances with microcidal, cytotoxic, cytolytic, apoptotic, anti-angiogenic, or anticoagulant qualities. This paper addresses the significance of bioactive compounds from insects as lead compounds for producing new therapeutics.
Key words
insects, bioactive compounds, novel structures, unique mechanisms, new drug discovery and development
Introduction
Today, approximately 80% of the human population living in developing countries still relies for an important part on naturally-derived substances, particularly plants with medicinal properties, for its day-to-day health care [1,2]. The remaining 20% of individuals in the industrialized parts of the world uses in 25% of cases drugs that have been directly derived from natural products [1,2]. This underscores the importance of the biodiversity to both traditional medicinal systems and modern, allopathic medicinal practices.
A myriad of breakthrough therapeutics has been developed by following up naturally-derived leads with medicinal properties, not only from plants but also from microorganisms and marine species [3,4]. A few examples are the cardiac glycoside digoxin from the foxglove Digitalis lantana L. (Scrophulariaceae) for treating certain heart conditions [5]; the muscle relaxant D-tubocurarine derived from the Amazon plants Strychnos toxifera L. (Loganiaceae) and Chondrodendron tomentosum L. (Menispermaceae) [6]; the hypoglycemic agent metformin synthesized on the basis of the chemical structure of galegine in the French lilac Galega officinalis L. (Fabaceae) [7]; the antibiotic penicillin from the fungus Penicillium chrysogenum Thom, 1910 (Trichocomaceae) [8]; the powerful pain killer ziconotide molded on the basis of conotoxin in the venom of the cone snails Conus geographicus Linnaeus, 1758, and Conus magus Linnaeus, 1758 (Conidae) [9]; and the novel antineoplastic agent trabectedin (Yondelis®) originally isolated from the Caribbean sea squirt Ecteinascidia turbinata Herdman, 1880 (Perophoridae) [10].
However, when compared to plants and marine species, the animal kingdom, particularly the order of the insects (Insecta Linnaeus, 1758), represents at least a similar abundance of bioactive compounds with potentially useful therapeutic properties. The insects are the most varied group of animals on our planet, comprising a biomass of a minimum of 2.6 million and a maximum of 7.8 million individuals [11,12] and have colonized a wide variety of terrestrial, freshwater, and even a few marine habitats [11,12]. Thus, when taking an average of 5.5 million insect individuals, this group of animals encompasses over 70% of the estimated 7.77 million extant animals on Earth [11,12].
Taxonomically, the Insecta represent a class of invertebrates within the Hexapoda, a subphylum of the Arthropoda ([11,12]; Box 1). The extinct Trilobitomorpha as well as the extant Chelicerata (e.g., spiders and scorpions), Myriapoda (e.g., centipedes and millipedes), and Crustacea (e.g., lobsters, crabs, and shrimps) also belong to the Hexapoda ([11,12]; Box 1). The insects comprise an estimated 950,000 different species classified in approximately thirty orders [11,12]. The orders of the Mantophasmatodea (gladiators), Grylloblattodea (icecrawlers), and Zoraptera (angel insects) only include twenty-four to thirty-two species [11,12]. On the other hand, highly species-rich orders such as the Orthoptera (e.g., grasshoppers, locusts, katydids, and crickets), Diptera (flies), Hemiptera (bugs), Hymenoptera (e.g., wasps, bees, ants, and sawflies), Lepidoptera (e.g., butterflies and moths), and Coleoptera (beetles) roughly consist of 25,000 to 400,000 different species [11,12].
Box 1. Taxonomic position of the insects in the animal kingdom (Based on Gullan PJ, Cranston PS, 2014)
Kingdom Animalia – animals
Phylum Arthropoda – arthropods
Subphylum Trilobitomorpha
Class Trilobita – trilobites (extinct)
Subphylum Chelicerata
Class Arachnida – spiders, scorpions, etc.
Class Merostomata – horseshoe crabs, eurypterids
Class Pycnogonida – sea spiders
Subphylum Myriapoda
Class Chilopoda – centipedes
Class Diplopoda – millipedes
Class Pauropoda – sister group to millipedes
Class Symphyla – resemble centipedes
Subphylum Crustacea
Class Branchiopoda – brine shrimp etc.
Class Remipedia – blind crustaceans
Class Cephalocarida – horseshoe shrimp
Class Maxillopoda – barnacles, copepods, fish lice, etc.
Class Ostracoda – seed shrimp
Class Malacostraca – lobsters, crabs, shrimp, etc.
Subphylum Hexapoda
Class Insecta – insects
Class Entognatha – springtails, etc.
Insects are characterized by a hard, jointed chitinous exoskeleton that encloses a body consisting in adults of three fused segments, viz. the head with a series of complex mouthparts and one pair of antennae, the thorax which carries three pairs of legs and usually two pairs of wings, and the abdomen which contains the digestive and reproductive organs ([11,12]; Figure 1). Both the antennae and mouthparts have probably evolved from modified walking legs, and the wings may be reduced or absent in a few very primitive species, becoming non-functional or adapted for a different purpose [11,12]. Notably, a number of other arthropods such as centipedes, millipedes, scorpions, and spiders also have a jointed skeleton (as holds true for all arthropods) but substantially differing features, most noticeably the absence of six legs in the adult stages [11,12].
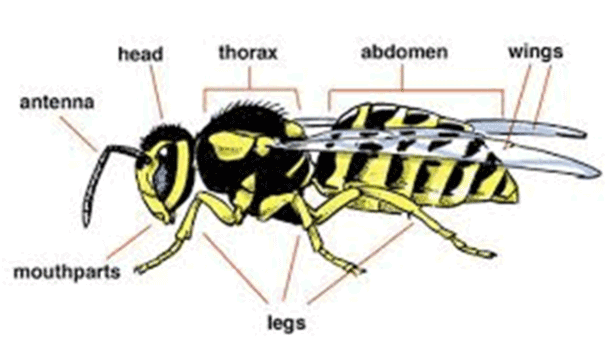
Figure 1. Basic body plan of the insects (from: http://www.earthlife.net/insects/anatomy.html)
Insects presumably evolved from a group of crustaceans [13] and first appeared in the Ordovician, some 480 million years ago [14]. The currently existing species can be classified into the subclasses Apterygota and Pterygota. The Apterygota are considered the most primitive insects alive today, and consist of at least four orders characterized by wingless adults (like the immature stages) which have not undergone a metamorphosis [11-14]. The Pterygota comprise winged and secondarily wingless insects, and can further be divided into at least sixteen orders within the Exopterygota and at least nine orders belonging to the Endopterygota [11-14]. The Exopterygota produce larvae that do not enter a pupal stage in their transition to adults, have nymphs that gradually develop into adults through moulting and already resemble adults, and develop wings outside their body [11-14]. The Endopterygota produce larvae that do not resemble adults, undergo an elaborate metamorphosis involving a pupal stage, and develop wings inside the body during pupation [11-14].
Background
Insect species communicate with each other and with their environment using a variety of tactile, visual, auditory, and chemical signals [15,16]. The chemical signals are referred to as semiochemicals or infochemicals and pass on information on, for example, impending threats and new food sources [15,16]. Important groups of insect chemicals are pheromones and allelochemicals [15,16]. Pheromones carry information from one individual to (an)other member(s) of the same species and include, among others, sex attractants, intraspecific aggregation signals, alarm substances, and trail-marking compounds [15,16]. Allelochemicals travel from one insect to (a) member(s) of (a) different species and include, for instance, defensive signals such as repellents and toxins as well as compounds to locate suitable host plants [15,16].
Many species of insects synthesize their own chemicals, but others derive these substances from the plants they feed on [15,16]. Some of these chemicals - such as alkaloids, bufadienolides, cantharidins, cyanides, and cardenolides - act as powerful defensive poisons that restrict movement of attackers [15,16]. Such compounds can irritate, injure, induce vomiting, cause blistering, or even kill the enemy [15,16]. Others function as repellents that stimulate scent and taste receptors of predators, discouraging them from feeding [15,16]. Such substances comprise in general volatile and reactive chemicals like acids, aldehydes, aromatic ketones, quinones, and terpenes [15,16]. Some insect species use combinations of defensive chemicals and repellents, and some chemicals can exert multiple effects [15,16].
For instance, when threatened, the soldiers of the pantropical termite subfamily Nasutitermitinae (Blattodea: Termitidae) exude noxious liquids containing several monoterpene hydrocarbons through a horn-like frontal projection [17]. The red spot assassin bug Platymeris rhadamanthus Gerstaecker, 1873 (Hemiptera: Reduviidae) immobilizes its prey by using venomous saliva that contains potent lytic enzymes which cause intense local pain, vasodilation, and edema, and help in the external digestion of prey [18]. Soldiers of the Malaysian carpenter ant Camponotus saundersi Emery, 1889 (Hymenoptera: Formicidae) violently release a sticky corrosive and irritant substance from their large mandibular glands running through their entire body, causing the body to explode, thereby engulfing the attacker with the poison [19]. And bombardier beetles in the tribes Brachinini Bonelli, 1810; Paussini Latreille, 1807; Ozaenini Hope, 1838; and Metriini LeConte, 1853 (Coleoptera: Carabidae) spray boiling hot, toxic, and foul-smelling quinonoids from their abdomen in the direction of a predator [20].
As well, larvae from the three-lined potato beetle Lema daturaphila Kogan & Goeden, 1970 (Coleoptera: Chrysomelidae) use poisonous fecal shields produced from their diet of toxic alkaloid-containing plants such as nightshades to deter predators [21]. The North American eastern lubber grasshopper Romalea microptera Houttuyn, 1813 (Orthoptera: Romaleidae) emits a foul-smelling and foul-tasting foamy mixture of phenols, terpenes, and benzoquinones from its thorax when disturbed [22]. And when brought into an anthill, caterpillars from the Japanese oakblue Arhopala japonica Murray, 1875 (Lepidoptera: Lycaenidae) secrete a sticky, sweet substance that presumably modifies brain dopamine signaling of the ants, manipulating the ants to protect and even pamper the caterpillars instead of eating them [23].
In light of their effectiveness as warning signals and anti-predator defenses, the meaningful pharmacological properties of certain insect chemicals should be beneficial against human diseases. The above-mentioned reduviid venom, for instance, may treat bacterial infections in humans [24]. Apitoxins such as melittin cause inflammation after bee stings, but these compounds and some of their analogues have been reported to induce immunomodulatory, cytotoxic, and apoptotic effects in various preclinical models [25]. And cantharidin, the defensive chemical from blister beetles, may represent the lead compound of a promising new class of antineoplastic agents [26].
These and many more examples underscore the pharmacological diversity of insect-derived bioactive substances and their potential therapeutic usefulness. This paper addresses a number of unique bioactive compounds from this group of animals, their (putative) mechanism(s) of action, as well as their potential clinical applicability, and closes with the previsions of new drug development programs exploring these substances. The compounds have been grouped on the basis of the insect order in which they have been encountered (Table 1). The insect orders have been arranged according to their approximate evolutionary complexity with the oldest and most primitive groups addressed first [13,14].
Table 1. Bioactive compounds with potential clinical applicability derived from insect species. The insect species have been grouped according to the taxonomic order to which they are assigned, and the insect orders have been arranged according to their approximate evolutionary complexity with the oldest and most primitive groups addressed first
(Infra)Order |
Species (Family) |
Bioactive compound |
(Potential) clinical application(s) |
Orthoptera |
Brachystola magna Girard, 1854 (Romaleidae) |
Isocarbostyril alkaloids |
Anticancer agents |
Isoptera |
Pseudocanthotermes spiniger Sjöstedt, 1900 and Nasutitermis corniger Motschulsky, 1855 (Termitidae) |
Termicin and spinigerin |
Antimicrobial and antiparasitic agents |
Coleoptera |
Lytta vesicatoria Linnaeus, 1758 (Meloidae) |
Cantharidin and derivatives |
Anticancer agents |
Diptera |
Simulium vittatum Latreille, 1802 (Simuliidae) |
Simukunin and analogues |
Anticoagulants |
Lepidoptera |
Pieris rapae Linnaeus, 1758 (Pieridea) |
Pierisins |
Anticancer agents |
Hymenoptera |
Apis mellifera Linnaeus 1758 (Apidae)
Lasioglossum laticeps Schenck, 1870 (Halictidae)
Vespula spp., Vespa spp., and Provespa spp. (Vespidae)
Solenopsis invicta Buren 1972 (Formicidae)
Pseudomyrmex triplarinus Weddell, 1850 (Formicidae) |
Melittin
Lassioglossins
Mastoparans
Solenopsin
Myrmexins |
Antimicrobial, anti-inflammatory, and anticancer agents
Antimicrobial and anticancer agents
Antimicrobial and anticancer agents
Anticancer agents
Rheumatoid arthritis |
Bioactive compounds from Orthoptera
The order Orthoptera includes, among others, grasshoppers, crickets, and locusts, and represents, with approximately 27,000 species in 235 subfamilies and families worldwide, one of the largest groups of insects [11,12]. Their name derives from the Greek words ‘ortho’ meaning ‘straight’ and ‘pteron’ meaning ‘wing’ [11,12]. These insects have a characteristically cylindrical body and strong, enlarged legs adapted for jumping, and many species communicate with one another by sounds produced by rubbing their wings against each other or against their legs [11,12].
Representing valuable sources of proteins, certain species of Orthoptera are part of the diets of various societies throughout the world [27]. Others are incorporated in the traditional medicinal practices of, for instance, indigenous communities in India and Brazil [27,28]. Notably, the native peoples of Mexico already used the femurs of grasshoppers for treating liver ailments, presumably because of their resemblance to human liver [29].
Orthoptera feed on a large variety of plants including economically valuable agricultural crops [30]. Particularly grasshoppers and locusts have been regarded as major pests since biblical times, regularly attacking fields of crops in swarms of millions of individuals which wipe out entire areas in a few hours [30]. On the other hand, many Orthoptera are preyed on by various microbial pathogens as well as a number of reptiles, birds, and small mammals, while certain parasitic dipteran and hymenopteran species lay their eggs into the eggs or cuticles of orthopterans [30].
As a result, the Orthoptera have developed a number of remarkable chemical defense mechanisms for their protection. For example, many grasshoppers repel predators by ‘tobacco spitting’, i.e., by regurgitating their irritant or toxic digestive fluids on the predator [31]. Katydids from the genus Eugaster Serville, 1838 (Tettigoniidae) autohemorrhage in response to visual stimuli and can accurately eject blood up to forty centimeters in the direction of an attacker [31,32]. Other orthopteran species fortify their blood cells or internal organs with distasteful or toxic allomones [33,34]. And some lubber grasshoppers (family Romaleidae) discharge a defensive secretion from paired metathoracic spiracles to their enemy [31,32]. Studies exploring the chemicals produced by Orthoptera have led to the identification of, among others, a number of isocarbostyril alkaloids with interesting bioactivity.
Isocarbostyril alkaloids
Isocarbostyrils are alkaloids that do not contain basic nitrogen atoms and are represented by hydroxylated benzophenanthridone or isoquinolinone types of structure [35,36]. They were first isolated from plants of the Amaryllidaceae family which have long been used for their medicinal and toxic properties [37]. Notably, already in ancient Greece, the oil from Amaryllidacea members such as Narcissus species was used for treating cancer [37]. Particularly the isocarbostyril alkaloids narciclasine, lycoricidine, and pancratistatin isolated from various Amaryllidaceae members [35] displayed intriguing antitumor activity in various in vitro and in vivo models [38-42].
These cytotoxic effects occurred at concentrations that did not affect the growth of normal human lung fibroblasts [43]. Narciclasine, for instance, accomplished a 50% reduction in the number of viable tumor cells after three days of treatment at mean concentrations of 0.03 μM compared to 7.5 μM for the fibroblasts [43]. Subsequent mechanistic studies indicated that these effects were attributable to the selective induction of internucleosomal DNA fragmentation and the activation of the initiator caspases -8 and -10 of the death receptor pathway in tumor cells but not in normal human fibroblasts [43-47]. These observations suggest that isocarbostyril alkaloids may represent a promising class of novel apoptosis-inducing anticancer agents.
As part of an effort to explore terrestrial arthropods as potential sources of novel and structurally unique anticancer drugs, the Texas grasshopper Brachystola magna Girard, 1854 (Romaleidae) (Figure 2) was also demonstrated to contain isocarbostyril alkaloids, albeit at levels lower than those found in the Amaryllidaceae [48]. The compounds isolated were pancratistatin, narciclasine, and umgeremine, and presumably act as defensive chemicals for the grasshopper [48]. Particularly the two former compounds displayed meaningful in vitro and in vivo antitumor effects, comparable to those previously noted for the plant-derived compounds [48], emphasizing their potential usefulness as novel anticancer agents.
There are no records to indicate that B. magna specifically or preferentially feeds on plants of the Amaryllidaceae family [49]. This suggests that this insect species produces isocarbostyrils through its own metabolism and, given its biomass, may represent a feasible alternative source of these promising substances. This is all the more important because so far these compounds cannot be synthesized by cost-effective processes [35]. At the same time, several research groups are directing their efforts to acquire new isocarbostyril derivatives or analogues for further preclinical anticancer testing [35]. Such compounds may not only emerge by modulating the structures of the currently available compounds, but also by further exploring orthopteran species for their ability to synthesize such compounds through modified or alternative biochemical pathways.
Bioactive compounds from Blattodea
The insect order Blattodea (Greek for ‘an insect that shuns the light’) includes approximately 3,000 species of termites in roughly 300 genera of nine families, besides an estimated 4,600 species of cockroaches in about 500 genera of seven families [11,12]. Previously, the termites were considered a separate order (Isoptera), but relatively recent genetic and molecular evidence strongly suggests a close relationship with the cockroaches, both cockroaches and termites having evolved from a common ancestor [11,12]. For this reason, the termites have been classified as the infraorder Isoptera residing under the order of the Blattodea [11,12].
The Isoptera comprises approximately 3,000 termite species in roughly 300 genera of nine families [11,12]. The name of this infraorder is derived from the Greek words ‘iso’ and ‘pteron’ meaning ‘equal’ and ‘wings’, respectively, in reference to the similar size, shape, and venation of the four wings [11,12]. Termites are sometimes referred to as white ants but they are not related to this insect family, even though their colonies, like those of ants (as well as those of bees and wasps), comprise several castes including queens, workers, and soldiers [11,12]. They are detritivores, feeding on dead plant material and cellulose, and are encountered on all continents except Antarctica [11,12]. Their colonies can consist of several millions of individuals, and their queens may live up to fifty years [11,12].
Several species of termites - for instance, the Termitidae members Globitermes sulphureus Haviland and Neocapriterme taracua, as well as members of the family Serritermitidae - have developed dramatic ways to protect their colonies from intruders: they engage in self-destruction while releasing incapacitating and/or toxic compounds on their attackers [50-53]. In addition, presumably as a result of their feeding habits, many termite species produce substances with potent and broad antimicrobial activity that they store in various parts of their gastrointestinal tract [54]. Some of these compounds may represent interesting candidates for developing novel drugs against microbial and parasitic infections.
Termicin and spinigerin
Termite products also have a long use in several traditional medicinal systems, for instance, as a paste consisting of a mixture of macerated termites and parts of the mound [27,55]. In several traditional African medicinal systems, the paste is boiled and topically applied to prevent infection of external wounds, and ingested to treat internal hemorrhages [27]. And in Indian Ayurveda, the boiled paste is used as a remedy for ulcers, rheumatic diseases, anemia, pain, and a declining health [27,55]. Interestingly, termites are also used as a medical device in certain African countries. In order to insert a drug substance subcutaneously, a termite is allowed to bite into the area of the patient’s skin coated with the substance, thus effectively injecting it under the skin [55].
Relatively recent insights have made clear that the therapeutic efficacy of at least some of these traditional uses may be attributed to the antibiotics the insects produce [56,57]. Among these are the antimicrobial peptides termicin and spinigerin in the salivary glands of the Termitidae Pseudocanthotermes spiniger Sjöstedt, 1900 [58] and Nasutitermis corniger Motschulsky, 1855 (Figure 3) [59]. Their natural function is probably to kill a host of microorganisms and parasites [54,60], among others, by disrupting their cell membrane [60].

Figure 2. The Texas grasshopper Brachystola magna Girard, 1854 (Orthoptera: Romaleidae). From: http://www.naturalista.mx/observations/1921224

Figure 3. The cone head termite Nasutitermes corniger Motschulsky, 1855 (Blattodea: Termitidae). From: http://toutunmondedansmonjardin.perso.neuf.fr/EN/pages_EN/nasutitermes_sp._EN.htm
Not surprisingly, termicin displayed notable in vitro activity against several species of fungi as well as various Gram-positive and Gram-negative bacteria [58]. Notably, a decoction of the termite species N. corniger acted synergistically with the antibiotics neomycin, gentamicin, and erythromycin against cultured Escherichia coli and Staphylococcus aureus bacteria including multidrug resistant strains [58,61,62], presumably by inhibiting the drug efflux pump in the plasma membrane of the cells [59]. Moreover, spinigerin reportedly induced apoptotic-like cell death via a caspase-independent pathway in Leishmania (Leishmania) donovani parasites without affecting the host macrophages [63].
Together, these observations suggest that termite products - including termicin and spinigerin - may represent unprecedented modalities to overcome bacterial resistance to antibiotics and to fight parasitic infections such as visceral leishmaniasis. Thus, detailed evaluation of these compounds for their potential use against these conditions is warranted.
Bioactive compounds from Coleoptera
The insect order Coleoptera encompasses roughly 400,000 different species of beetles divided in four suborders and about 500 families and subfamilies [11,12]. The name of this group of insects is based on their sheathed wings, deriving from the Greek words ‘koleos’ for ‘sheath’ and ‘pteron’ for ‘wing’ [11,12]. The Coleoptera are the largest order in the animal kingdom, comprising approximately 25% of all known life forms and about 40% of all described insect species [11,12]. They characteristically exhibit a particularly hard exoskeleton and hard forewings [11,12]. The exoskeleton is made up of many plates joined by thin sutures, thus providing flexibility to the animal [11,12].
With the exception of oceans and arctic regions, the Coleoptera have colonized all types of habitats on Earth [11,12]. They often feed on fungi, organic debris, and other invertebrates, but also on plants including agricultural crops [11,12]. The Colorado potato beetle Leptinotarsa decemlineata Say, 1824 (Chrysomelidae) and the boll weevil Anthonomus grandis Boherman, 1843 (Curculionidae), for instance, are infamous as major agricultural pests in the Americas [64]. On the other hand, various ladybird beetles (family Coccinellidae) prey on agricultural pests such as aphids, scale insects, and thrips and are considered useful [11,12].
In their turn, many Coleoptera are prey for various birds and mammals [11,12]. As a result, they have developed a broad range of defensive chemicals to make themselves unpalatable or even toxic to predators [15,16]. Some of these substances are products from their own metabolism while others are secondary metabolites of the plants they feed on [15,16]. The toxic substances may be released by remarkable techniques, such as the jets of formic acid and the earlier mentioned boiling hot toxic quinonoids which are sprayed on potential predators from far distances [20].
Less spectacular but equally effective are the irritating and/or blistering chemical defenses of many Coleoptera such as cantharidin [66]. This substance displayed unique pharmacological properties, making it a subject of various new drug discovery and development programs [67]. The same holds true for some of its naturally occurring derivatives and chemically modified analogues like cantharimides [67-69], cantharidinimide [70,71], palasonin [71], and the demethylated cantharidin analogue norcantharidin [72].
Cantharidin and derivatives
Cantharidin is a terpenoid present in the dried and ground bodies of several species of blister beetles of the genera Mylabris and Lytta (Meloidae), especially the emerald-green L. vesicatoria Linnaeus, 1758 (Figure 4) [67,73]. It is produced by the male who gives it to the female during mating to cover the eggs as a defense against predators [67]. Also known as Spanish fly, cantharidin has a long use as an aphrodisiac in many societies [67,73]. However, the topical use or ingestion of this compound carries a substantial risk of causing toxicity including priapism and even death [67,73].

Figure 4. The blister beetle Lytta vesicatoria Linnaeus, 1758 (Coleoptera: Meloidae). From: http://www.terrarium.com.pl/326-zwierzeta-jadowite-i-trujace-w-otoczeniu-czlowieka/
Topical cantharidin has also been used for treating various skin disease such as furuncles and warts in particularly traditional Chinese and Vietnamese medicine [74], and since the 1950s in the USA for treating warts and molluscum contagiosum, a common viral disease of childhood caused by a poxvirus that presents with small, firm, dome-shaped, umbilicated papules [75]. This has led to its current approval of the US Food and Drug Administration (FDA) for “topical use in the professional office setting only” against warts and other skin problems [76].
More recent cell culture and animal studies indicated that catharidin effectively inhibited the growth of various human cancer cells in vitro and in vivo [77-79]. These findings might be attributable, at least in part, to the induction of apoptosis through the p53-pathway, causing oxidative stress and DNA damage [80-83]. This is followed by activation of the caspase-9 and caspase-3 pathways involving the mitochondrial intrinsic route [80-83], and activation of the JNK pathway subsequent to G2/M cell cycle arrest [84], respectively.
A number of studies suggest that the tumor growth inhibitory properties of cantharidins could be attributed, at least in part, to anti-angiogenic activity. Cantharidin and some of its derivatives inhibited the proliferation, migration, and invasion of, as well as capillary-like structure formation by cultured endothelial cells [85-88], as well as tumor growth and angiogenesis in various xenograft-mouse models [85-89]. Furthermore, norcantharidin treatment interfered with signaling for vascular development, presumably by inhibiting phosphorylation of critical elements of these pathways [86,91,92]. This led, among others, to a decreased microvessel density and a decreased expression of pro-angiogenic factors such as angiopoietin-2 [88] and upregulation of anti-angiogenic factors such as thrombospondin and tissue inhibitor of metalloproteinase [89]. Notably, norcantharidin also inhibited vasculogenic mimicry [89,90], an important cause of failure of treatment with anti-angiogenic anticancer agents.
The mechanisms underlying these observations probably include, at least in part, potent and selective inhibition of serine/threonine protein phosphatase activities, in particular those of type 1 protein phosphatase and type 2a protein phosphatase [67,93,94]. Together with protein kinases, protein phosphatases act as modulators of many intracellular signaling pathways including those involved in cell cycle progression, apoptosis, cell growth, and vascular development, operating as major ‘on’ and ‘off’ switches, respectively, in these pathways [77,84,85]. Clearly, either directly or indirectly manipulating the ‘off’ switches represented by protein phosphatases may represent an attractive strategy for controlling cancer. In this respect, particularly protein phosphatase 2a has been identified as a tumor suppressor for mainly blood cancers [95]. Thus, catharidin as well as some of its derivatives may represent very promising lead compounds for a novel class of antineoplastic agents.
Clinical evaluation of cantharidin as an anti-cancer agent was hampered by its relatively short half-life and high toxicity - mostly to kidneys and bone marrow [67] - as well as its poor water-solubility and low oral bioavailability [67]. The problem of the poor water-solubility has been solved by formulating cantharidin as a sodium salt (commercially known as Qinin®) that has mainly been evaluated in China for its efficacy against various disease conditons including solid tumors [96]. In one such a study, the addition of Qinin® to chemotherapy did not produce more responses in a cohort of seventy gastric cancer patients, but caused significantly less chemotherapy-related side-effects [97]. The problems of low bioavailability and substantial toxicity may be tackled by delivering cantharidin encapsulated by solid lipid nanoparticles [98] which would carry the drug close to the target, improving its antitumor efficacy and causing less toxicity to the normal tissues.
At the same time, other bioactive substances from blister beetles are being evaluated in a search for antitumor cantharidins with improved pharmacological properties over those of the parent compound. This has resulted in the isolation and identification of the cantharimides from the Chinese blister beetle M. phalerata Pallas (Meloidae) which exhibited protein phosphatase 1 and protein phosphatase 2a inhibitory activities similarly to those of cantharidin [67,68,78,99]. Also, structural modifications of cantharidin have led to several cantharidinimides with retained anticancer activity but less toxicity to non-cancer cells [99].
Particularly norcantharidin displayed interesting features besides countering angiogenesis [87,91,92]: it targeted cancer cells selectively while evoking less toxicity than cantharidin [100]; it overcame multidrug resistance in cultured breast cancer cells by inhibiting sonic hedgehog signaling and P-glycoprotein expression [101-103]; it modulated cancer stem cell self-renewal pathways [104,105]; it acted chemopreventive [106,107]; and it reduced the side-effects of radiation [108,109]. These encouraging results continue to position the Coleoptera and their bioactive products high on the radar of pharmacologists, clinical chemists, molecular biologists, and chemical ecologists who are active in the area of new drug discovery and development.
Bioactive compounds from Diptera
The insect order of the Diptera comprises the so-called true flies and includes an estimated 120,000 described species divided over two suborders [11,12]. The Diptera include many familiar insects such as mosquitoes, midges, house flies, sand flies, gnats, and blow flies which are found in all terrestrial niches with the exception of the polar zones [11,12]. Their name derives from the Greek words ‘di’ for ‘two’, and ‘pteron’ for ‘wing’, as they only use the forewings to fly, the hindwings acting as balancing organs known as halteres. Other distinguishing features of this insect order are their mobile head with a pair of large compound eyes, and mouthparts designed for piercing and sucking or for lapping and sucking [11,12].
Certain dipteran species such as leaf miner flies (family Agromyzidae), fruit flies (families Tephritidae Newman, 1834 and Drosophilidae Róndani, 1856), and gall midges (family Cecidomylidae) are important agricultural pests, while others such as tsetse flies (family Glossinidae Theobald, 1903) attack livestock causing substantial economic harm [11,12]. Still others such as mosquitos (family Culicidae Meigen, 1818) act as vectors for many infectious diseases like malaria, dengue, chikungunya, zika, West Nile virus, yellow fever, and encephalitis [110]. On the other hand, the Diptera probably are the most important pollinators after the Hymenoptera, particularly in wet and cold environments [11,12].
Many adult dipteran females feed on vertebrate blood which they use as a source of protein for the production of their eggs [11,12]. For this purpose, they pierce the skin of their victim with their sharp stylet and insert a complex cocktail of salivary components into the wound including a host of antihemostatic factors such as anticoagulants, inhibitors of platelet aggregation, anti-inflammatory substances, immunomodulatory components, and vasodilators [111-115]. These substances facilitate blood feeding by helping to overcome host defenses against blood loss, inflammatory reactions, disruption of blood flow, and blood clotting at the feeding site [111-115].
As a result, Diptera have been used for centuries by practitioners of Eastern Medicine to prevent thrombosis [116]. Subsequent studies on these substances led to the identification of, among others, simukunin that may represent the lead compound of a new class of potent anticoagulants.
Simukunin and analogues
Blood coagulation is a complex process that occurs almost immediately following tissue injury, and that involves the aggregation of platelets to form a plug at the site of injury [117]. This is accomplished by the simultaneous activation of an enzymatic cascade primarily consisting of serine proteases that ultimately leads to the formation of fibrin strands which strengthen the platelet plug [117]. Briefly, tissue factor (TF), a protein present in sub-endothelial tissue and leukocytes, binds to the activated blood coagulation factor VII (FVIIa), after which the TF/FVIIa complex activates factor X (FX) to produce factor Xa (FXa) [117]. FXa then combines with activated factor V in the presence of calcium and phospholipids to produce thrombin from prothrombin [117]. Thrombin acts as a serine protease that, among others, converts soluble fibrinogen into insoluble strands of fibrin [117]. Negative regulators of coagulation are primarily protease inhibitors including tissue factor pathway inhibitor (TFPI) [118] that inhibits formation of FXa [118,119].
As mentioned above, blood-feeding arthropods also produce anti-hemostatic factors in their saliva which facilitate blood feeding by interfering with host hemostatic responses [111-115]. A variety of anti-coagulation factors has been isolated from arthropods including the 140- and 308-amino acid proteins ixolaris and penthalaris, respectively, identified in the salivary glands of the deer tick Ixodes scapularis Say, 1821 (Ixodida: Ixodidae) [119,120], a vector of Lyme disease [120,121]. These proteins tightly bind to both FX and FXa, strongly inhibiting the TF/FVIIa complex and thus blood clotting [119,120].
Potent anticoagulation factors with activity against thrombin, FXa, and FV were also encountered in the salivary glands of females of the blood-feeding black fly Simulium vittatum Latreille, 1802 (Simuliidae) (Figure 5) [122-125]. S. vittatum is an important vector of the nematode Onchocerca volvulus Bickel, 1982 (Onchocercidae), the causative agent of onchocerciasis or river blindness [126]. One of the anticoagulants identified in the fly’s saliva was the polypeptide simukunin [127] that effectively blocked the blood coagulation cascade by inhibiting FXa [128]. Simukunin seems as effective as heparin in preventing the formation of blood clots, but unlike heparin, probably does not cause allergic reactions or thrombocytopenia triggered by immunological attack on platelets [128]. Importantly, this substance also inhibited the serine proteases elastase and cathepsin G and displayed encouraging anti-inflammatory properties [128], providing additional benefits over heparin with respect to antagonizing clot formation and limiting tissue damage caused by inflammatory responses.

Figure 5. The black fly Simulium vittatum Latreille, 1802 (Diptera: Simuliidae). From: http://bugguide.net/node/view/624421/bgpage
Other anticoagulants from the salivary glands of hematophagous insects with potential therapeutic value are tablysin-15, anophelin, and nitrophorin-2. Tablysin-15 is produced by the horsefly Tabanus yao Macquart, 1855 (Diptera: Tabanidae) - a vector for filariasis [129] - and inhibits platelet aggregation by interfering with the function of integrins on the platelet membrane [130]. Anophelin is a 6342.4-Da peptide identified in the Central American and African dipterans Anopheles albimanus C.R.G. Wiedemann, 1820 (Culicidae) [131] and A. gambiae Giles, 1902 [132], respectively, that tightly binds thrombin [131,132]. Nitrophorin-2 (or prolixin S) is a 20-kDa lipocalin in the kissing bug Rhodnius prolixus Stål, 1859 (Hemiptera: Reduviidae) [133] that associates with high affinity to either FIX or FIXa [134,135] and generates nitric oxide that causes vasodilatation and inhibition of platelet aggregation at the feeding site [136,137].
Clearly, advances in our understanding of the chemical structures and mechanisms of the components in the salivary glands of arthropods including d dipterans, may help in developing novel forms of treatment of conditions requiring the use of anticoagulants such as strokes, cardiovascular blockages, deep vein thrombosis, and pulmonary embolism [117]. In the meantime, ixolaris was found to block intracellular signaling pathways for pro-angiogenic substances in several human carcinoma cell lines while inhibiting the proliferation and metastasis of, and the vascularization of tumor cell grafts in nude mice [138-140]. Apparently, ixolaris also possesses meaningful anti-angiogenic qualities besides useful anticoagulant properties. Whether this also holds true for the substances in the salivary glands of the dipteran species mentioned above - including those of S. vittatum - must be verified in future studies.
Bioactive compounds from Lepidoptera
The insect order Lepidoptera consists of about 180,000 different species in 126 families and forty-six superfamilies, and includes moths and butterflies [11,12]. This group of insects comprises roughly 10% of the total number of described species on Earth [11,12] and is, together with the Diptera, Coleoptera, and Hymenoptera, among the largest orders of organisms on our planet [11,12]. The name ‘Lepidoptera’ is derived from the Greek words ‘lepidos’ for ‘scale’ and ‘pteron’ for ‘wing’, and refers to the scales covering particularly the wings of these insects, giving them their extraordinary variety of colors and patterns [11,12]. The colors and patterns help the animals with, among others, camouflage and mimicry, and act as signals to rivals and potential mates [11,12].
The Lepidoptera are found on all continents except Antarctica, mostly inhabiting terrestrial habitats associated with flowering plants [11,12]. In these ecosystems, many lepidopteran species play vital roles as pollinators [11,12]. This has led to the practice of butterfly rearing, the commercial breeding of butterflies to pollinate crops in greenhouses [141]. On the other hand, moths and butterflies are preyed on by, among others, birds, small mammals, and reptiles [11,12]. Particularly the larvae - caterpillars hatching from eggs laid near or on host plants, often causing considerable damage to agricultural crops [142] - are easy prey to assailants [11,12]. They defend themselves by camouflage or mimicry, but also by producing noxious and toxic substances, sometimes from secondary metabolites from the plants they feed on [15,16]. Some of these chemicals possess interesing pharmacological properties.
Pierisins
Similarly to many other insect species, lepidopterans are since long used in various traditional medicinal systems. The indigenous Central American peoples, for instance, boiled pupae of the silkworm - the larvae of the silk moth Bombyx mori Linnaeus, 1758 (Bombycidae) - to treat apoplexy, aphasy, bronchitis, pneumonia, convulsions, hemorrhages, and polyuria [29]. And in Indian Ayurveda, the larvae from the Jatropha leaf miner Stomphastis thraustica Meyrick, 1908 (Gracillariidae) - a major pest to plant species belonging to the genus Jatropha L. (Euphorbiaceae) [143] - are boiled and ground to give a paste that is topically applied to induce lactation, reduce fever, and relax an upset gastrointestinal tract [55].
More recently, a series of proteins designated pierisin-1a and -1b as well as pierisin-2 through -5 has been identified in both adults and pupae of butterfly species of the genus Pieris (Pieridea) including the cabbage butterfly P. rapae Linnaeus, 1758 (Figure 6) [144-147]. The detrimental effects of pierisin-1 on eggs and larvae of endoparasitic wasps that insert their eggs inside P. rapae pupae [148] suggest that these compounds function as chemical defenses against microbes and/or parasitoids.

Figure 6. The cabbage butterfly Pieris rapae Linnaeus, 1758 (Lepidoptera: Pieridea). From: http://www.amateuranthecologist.com/2015/12/a-year-of-pollinators-moths-and.html
Subsequent in vitro studies indicated that pierisins -1 and -2 elicited meaningful cytotoxic and apoptotic effects in various human carcinoma cell lines [144,147,149-152] and suppressed the growth of HeLa cells engrafted in BALB/c nude mice [153]. The induction of apoptosis occurred through a mitochondrial pathway involving Bcl-2 and caspases [152,153].
Notably, pierisin-1 mRNA expression increased during maturation of the pupae and was highly expressed in late-stage pupae, and its protein product accumulated in fat bodies where it persisted during pupation [154,155]. These observations suggest that pierisin-1 plays a critical role during metamorphosis of the pupae by removing unnecessary cells. Thus transforming P. rapae pupae into adults, providing support for its presumed ability to cause apoptosis.
Other studies indicated that each pierisin isoform contains a mono(ADP-ribosyl)transferase enzyme domain and receptor-binding domains [146] and that these compounds were able to catalyze the covalent transfer of ADP-ribose moieties from NAD to the N-2 position of guanine bases in DNA [151]. This resulted in the generation of more than 106 ADP-ribosylated DNA adducts per molecule of pierisin-1 [154]. These observations suggest that the cytotoxic and apoptotic actions of pierisins are mediated by the formation of guanine-specific ADP-ribosylated DNA adducts.
Taken together, pierisins and their derivatives, first described as novel DNA-modifying toxins, may play important roles in the treatment of cancer through the activation of apoptosis following the formation of adducts with DNA.
Bioactive compounds from Hymenoptera
The order Hymenoptera comprises the third largest group of insects, consisting of over 150,000 described species including ants, bees, wasps (including hornets), and sawflies [11,12]. The name of the order derives from the Greek words ‘hymen’ and ‘pteron’, signifying a key characteristic of this group of insects, viz. the connected hind and fore membranous wings [11,12]. Another morphological feature typical for Hymenoptera - at least in females - is the presence of an ovipositor for inserting eggs into hosts or inaccessible places [11,12]. In many species, the ovipositor is modified to a stinger, and the eggs are laid from the base rather than from the tip which is only used to inject venom to immobilize prey or in defense [11,12].
With the exception of extreme cold, remote, and inhospitable locations, hymenopterans have colonized all terrestrial habitats on Earth [11,12]. As a group, these insects have a wide range of feeding habits. Some are herbivorous, mainly feeding on leaves; bees feed on nectar and pollen; stinging wasps provide their larvae immobilized prey; and the larvae of certain species are parasitoids, feeding on a paralyzed prey after the eggs have hatched inside the victim [11,12].
The venoms from many hymenopteran species comprise a source of numerous bioactive compounds which evolved for prey capture and defense against microorganisms, rival colonies, and predators [157]. The venoms often consist of complex mixtures of bioactive amines including serotonin, histamine, tyramine, dopamine, noradrenaline, and adrenaline; pain-producing peptides such as kinins; and many types of hydrolases such as proteases, hyaluronidases, phosphatases, nucleotidases, and phospholipase A; as well as allergens and neurotoxins [156].
Many ant species produce these compounds in their metapleural glands on their thorax and use them as broad-spectral antimicrobial agents to prevent infections within the colonies [157]. The venoms of most honey bees and wasps (including hornets) also contain antimicrobial peptides [158-160]. Another chemical defense of Hymenoptera includes the venom of the honey bee Apis mellifera Linnaeus, 1758 (Apidae) (Figure 7) [161] which contains melittin, a powerful detergent that provokes hemolysis of red blood cells [162]. And wasps (subfamily Vespinae) use their venoms to paralyze and kill prey or defend the nest [163].

Figure 7. The honey bee Apis mellifera Linnaeus, 1758 (Hymenoptera: Apidae). From: http://www.insectimages.org/browse/detail.cfm?imgnum=2116051
When considering the potency of hymenopteran venoms and their relatively high selectivity to their molecular target, it is understandable that these substances have been used medicinally for centuries in various cultures. For instance, traditional Chinese medicine recommends the consumption of the edible black ant Polyrhachis vicina Roger, 1863 (Formicidae) to increase virility and fertility, prolong life, delay aging, and enhance mental performance [164]. More recent studies showed encouraging analgesic and anti-inflammatory activities by extracts and fractions from this hymenopteran [165]. Furthermore, the indigenous Central American peoples induced an immunological reaction via direct stings by the red harvester ant Pogonomyrmex barbatus Smith, 1858 (Formicidae) - that causes local inflammation and acts as an anticoagulant [29] - to fight arthritis, rheumatism, polyneuritis, poliomyelitis, and asthma [29]. They also used army ant soldiers (family Formicidae) as stitches to hold together the edges of a wound [29]. For this purpose, the ants were allowed to bite into the wound edges after which their thorax and abdomen were removed, leaving the heads holding the edges together [29]. Later, the antibiotics present in the ants’ salivary gland secretions were found to aid in the healing process [29].
Thus, bioactive compounds from the venoms of bees, wasps and ants represent promising lead compounds for developing novel therapeutics against human diseases. Some of these compounds include melittin and lasioglossins from bee species; mastoparan from wasp species; and solenopsin A and analogues as well as myrmexins from ant species.
Melittin
Products from A. mellifera such as honey, propolis, royal jelly, and bee bread have been used for centuries in apitherapy on every continent of the world [29,166]. The same holds true for apitoxin (bee venom), a complex mixture of several biologically active proteins and neurotransmitters such as phospholipases A2 and B, hyaluronidase, serotonin, histamine, dopamine, noradrenaline, and adrenaline, some of which can contribute to the clinical signs and symptoms of envenomation [167]. The beneficial properties of apitoxin and several of its components are since long known and have been widely used in Oriental traditional medicine to relieve pain, and to treat inflammatory diseases such as rheumatoid arthritis, multiple sclerosis, and tendonitis [168]. For these purposes, the bee venom was administered by live stings, injection of venom, and venom acupuncture [168].
Apitoxin has also been used for treating cancer [169], skin conditions [170] and even Parkinson’s disease [171]. In addition, a purified bee venom preparation called Apitox® has been approved by the FDA as a subcutaneously injectable product for relieving pain and swelling associated with rheumatoid arthritis, tendinitis, bursitis, and multiple sclerosis [172]. The results from a number of clinical studies provided some support for this decision [173,174].
One of the principal components of bee venom is melittin, an amphiphilic linear peptide of twenty-six amino acids with an amino-terminal region that is predominantly hydrophobic and a carboxyl-terminal region that is hydrophilic [175]. As a result, it is water-soluble but yet can spontaneously associate with natural and artificial membranes [176,178]. This feature enables melittin to accumulate in cell membranes, disrupting their phospholipid backbones and causing cell lysis [162,175,177]. Of note, melittin not only induces the lysis of a wide range of plasmatic membranes but also of intracellular membranes such as those of mitochondria [176-178]. Melittin is a powerful stimulator of phospholipase A2 that is also present in bee venom, and the synergistic action of these two substances enforces the cytolytic and cytotoxic effects started by the former compound [189].
These events may lead to the release of other harmful compounds from the damaged cells such as lysosomal enzymes from leukocytes, serotonin from thrombocytes, and histamine from mast cells [167]. However, a large body of preclinical studies also suggest that melittin may elicit multiple pharmacological effects, including antibacterial actions [180,181], antiviral properties [182,183], inhibition of cancer cell growth [184,185], inhibition of cancer cell metastasis [186], interference with angiogenesis [184,187,188], apoptosis-inducing effects [189], radiosensitization [190], immunosuppression [191], anti-inflammatory effects [192,193], as well as anti-atherosclerotic effects [194,195].
The precise mechanisms underlying all these actions remain to be elucidated. However, the antibacterial effects and potential anticancer efficacy of this membrane-active peptide may be related to cytolysis following the activation of phospholipase A2, the induction of pores or perturbations in plasma and subcellular membranes, and the activation of apoptotic pathways such as those mediated by caspases and metalloproteinases [25,162,196]. On the other hand, its presumed ability to inhibit phospholipase A2 [192] suggests that mellitin may be useful against inflammatory, rheumatic, and atherosclerotic conditions.
Despite encouraging laboratory data, so far no offically approved medical products for human use that are based on mellitin are on the market. Various studies have focused on applications in the field of bacterial and viral infections, inflammatory conditions such as rheumatoid arthritis and arteriosclerosis, and cancer [162,196]. Although the conversion of all these possibilities into therapeutic applications is still far away, these observations underscore the therapeutic prospectives of mellitin.
Lasioglossins
Lasioglossins represent a series of three structurally related pentadecapeptides in the venom of the eusocial bee Lasioglossum laticeps Schenck, 1870 (Halictidae) (Figure 8) [197]. Lasioglossin I adopted an α-helical structure in a membrane-mimetic environment of dodecylphosphocholine micelles, the polar side chains aligning along one side and the hydrophobic residues along the opposite side of the helical coat [198]. Thus, lasioglossins are membrane-active amphiphilic structures which, comparably to mellitin [162,175-178], have the capacity to adsorb to, insert into, readily cross cell membrane bilayers to interact with cytoplasmic constituents [198].

Figure 8. The wild bee Lasioglossum laticeps Schenck, 1870 (Hymenoptera: Halictidae). From: http://www.naturspaziergang.de/Bienen-2.htm
The result is perturbation of biological membranes in a way comparable to mellitin [198,199]. In addition to causing membrane destabilization, lasioglossins may tightly bind to DNA [198]. These actions may be responsible for their strong antimicrobial activity against both Gram-positive and Gram-negative bacteria without causing substantial hemolysis and mast cell degranulation, in addition to cytotoxic activity against various cancer cell lines [197]. Subsequent studies showed that the lasioglossins also displayed encouraging and prompt in vitro activity against several Candida species [199].
These observations suggest that lasioglossins may represent promising leads for the development of new, effective drugs against bacterial and fungal infections and possibly also against cancer. Their uncommon mechanism of action - perturbation of the structure of membranes and perhaps also that of DNA - may prevent the emergence of resistant microbial strains (and perhaps also that of resistant cancer cells).
Mastoparans
Mastoparans are tetradecapeptides in the venoms of vespid wasps (family Vespidae) [200] and several species of hornets (insects in the genera Vespa and Provespa residing under the Vespidae) (Figure 9) [201]. Like mellitin and lasioglossins, mastoparans are characterized by a hydrophobic and a hydrophilic domain [202] which provides them, similarly to the former compounds, amphiphathic features and the ability to disrupt or cause pores in biological membranes [203,204].

Figure 9. The wasp Vespula spp. (Hymenoptera: Vespidae). From: http://www.capetowninvasives.org.za/project/animals/species/vespula-germanica
Extensive research over recent decades has shown that mastoparans possess meaningful pharmacological properties [163] including cytolysis following destabilizing of plasma membranes [205]; perturbation of transmembrane signaling by directly interacting with G proteins [203,206-208]; the induction of apoptosis subsequent to the stimulation of phospholipases and mobilization of Ca2+ from mitochondria and sarcoplasmic reticulum [209-212].
These properties are probably involved in the antimicrobial studies and antiviral effects of mastoparans seen in preclinical [213-216]. Mastoparans displayed, in addition, noteworthy antitumor activity in vitro [203,218,219] and in vivo [220]. The main mechanism presumed to underlie these cytotoxic effects is mitochondrial permeability transition initiated by opening of high conductance permeability transition pores in mitochondria, leading to necrosis and apoptosis [209-212]. As a result, however, the toxicities caused by these compounds when used against cancer, may particularly involve mitochondrial permeability transition in normal tissues [221]. Should this holds true, these side-effects may be mimimized by more specifically delivering mastoparans to tumor cell mitochondria encapsulated in a liposomal preparation [221].
Meanwhile, several mastoparan analogues with retained potency but improved pharmacological features have been produced. Examples are the mitoparans which possess greater amphiphilicity when compared to the mastoparans [222], as well as the capacity to selectively bind tumor cell adhesion molecules [223]. Furthermore, a number of other mastoparan peptides called polybia-MPI, polybia-MPII, and polybia-MPIII has been identified in the venom of the Brazilian wasp Polybia paulista Ihering, 1896 (Vespidae) [224,225]. Polybia-MPI suppressed the growth of cultured prostate and bladder cancer cells as well as that of multidrug resistant leukemia cells by producing pores in the plasma membranes following interaction with membrane phospholipids [226]. Importantly, this compound induced substantially less cytotoxicity to normal murine fibroblasts [226], suggesting that it had some selectivity for malignant cells. Polybia-MPII and polybia-MPIII caused pronounced lysis of rat mast cells and erythrocytes and displayed antimicrobial activity against both Gram-positive and Gram-negative bacteria [228].
These findings are encouraging and suggest that the mechanistically unique mastoparans will one day be incorporated into every-day clinical practice for treating, among others, microbial infections and cancer.
Myrmexins
The tropical devil-tree ant Pseudomyrmex triplarinus Weddell, 1850 (Formicidae) (Figure 10) lives in symbiosis with trees of the South American genus Triplaris and protects them from predation by other insects and animals [228]. The ants are very territorial and their sting is extremely painful [229]. The venom has intense phospholipase activity and intermediate hemolytic activity [229]. According to folklore, certain Amazonian tribes punished members who had broken the rules by tying them to a Triplaris tree, exposing them to hundreds of ants’ stings until they begged for forgiveness for their mistakes [28]. As well, the indigenous people of the Peruvian Amazon treated patients suffering from rheumatoid arthritis by having the afflicted individuals bitten by the ants [28,29].

Figure 10. Group of tropical devil-tree ants Pseudomyrmex triplarinus Weddell, 1850 (Hymenoptera: Formicidae). From: https://goo.gl/images/sViBCy
A relatively recent analysis of the venom of P. triplarinus showed that it contained at least twelve proteins including phospholipase, hemolysins, as well as a multi-protein complex designated myrmexins I through VI [230]. Each of the six protein isoforms has a molecular mass around 7,000 Da, and each comprises a heterodimer consisting of a small subunit connected by a disulfide linkage to a larger, structurally unrelated subunit [230]. So far, the biological function(s) of the myrmexins remain(s) unknown.
However, both preclinical [231,232] and a double-blind, controlled clinical study [232,233] have shown that a partially purified extract of the venom from Pseudomyrmex spp. decreases pain and inflammation in patients with rheumatoid arthritis and reduces swelling in animal models of inflammation. Further investigations should clarify whether these compounds are indeed useful for treating rheumatoid arthritis and which myrmexin isoform is best suited as a remedy for this condition.
Solenopsins
Solenopsin A is a piperidine alkaloid in the venom of the so-called red imported fire ant Solenopsis invicta Buren, 1972 (Formicidae) (Figure 11). S. invicta presumably originates from Brazil and is a major threat to agricultural crops and livestock in various parts of the world [234]. This compound was identified in the 1930s in south-eastern USA during a project aimed at finding ways to exterminate this invasive ant species [234]. Analyses of its venom led in 1979 to the isolation of a number of toxic proteins and piperidines including trans-2-methyl-6-n-undecylpiperidine or solenopsin A [235]. At least some of these substances could be related to the various pharmacological properties of Solenopsis venom, including cytotoxic, hemolytic, necrotic, insecticidal, antibacterial, antifungal, anti-HIV, cardiodepressant, and neurologic actions [236].

Figure 11. The red imported fire ant Solenopsis invicta Buren, 1972 (Hymenoptera: Formicidae). From: https://goo.gl/images/stX3bM
More recent studies [237] showed that solenopsin A potently inhibited the growth of SVR cells (a transformed murine endothelial cell line) by inhibiting endothelial-specific signaling. This suggested that it has anti-angiogenic activity, which was corroborated by the delayed sprouting of intersomitic vessels from the dorsal aorta it caused in zebrafish [237]. Comparable results were found with solenopsin A and/or its analogues in the p53-deficient renal cell carcinoma cell line 786-O [237] and in various human melanoma cell lines, primary human melanocytes, as well as primary and immortalized human keratinocytes [238].
Studies with mouse embryonic fibroblast cell lines suggested that these effects occurred through the selective inhibition of a series of kinases involved in angiogenesis, most notably phosphatidylinositol-3-kinase (PI3K) and its downstream effector Akt [237,239,240]. These signaling elements play major regulatory roles in the control of apoptosis, proliferation, and angiogenesis [240-242], and are amplified or overexpressed in a number of malignancies [240,241, 243]. The PI3K/Akt pathway regulates the production of the potent angiogenic factor VEGF [240,242], and protects tumor cells against both chemotherapy and reactive oxygen-induced apoptosis [239,240,244].
When considering the selective targeting of Akt by solenopsin and its analogues, these compounds may have potential utility as anti-angiogenic antineoplastic agents. In fact, various inhibitors of PI3K/Akt such as the miltefosin analogue perifosine have undergone clinical testing in patients with advanced forms of cancer [245]. However, unsatisfactory response rates in several phase III studies led to the discontinuation of perifosine as an antineoplastic agent [246]. These disappointing results with perifosine emphasize the importance of solenopsin and its analogues as potentially novel Akt inhibitors for anti-angiogenic anticancer therapy.
Concluding remarks
This paper has reviewed the literature on the potential clinical applicability of a number of bioactive compounds from medicinal insects. Although only very few of these products have reached the clinic today, the results obtained so far are encouraging. Isocarbostyril alkaloids from grasshoppers, cantharidin and its derivatives from blister beetles, pierisins from butterfly larvae, as well as solenopsin A and its analogues from ant species may represent lead compounds of entirely new classes of anticancer agents [48,78,79,67,99-101,144,149,237,238]. Termicin and spinigerin from termite species, melittin and lasioglossins from bee species, as well as mastoparans from wasp species may help fight microbial and parasitic infections more effectively [58,59,62,64,180-183,197,199,213,216]. Simukunin and its analogues from certain blood-feeding insect species show us a way to develop unprecedented anticoagulants [127,128]. And myrmexins from ant species possess encouraging anti-inflammatory properties that are much needed in the clinic [231-233].
Admittedly, these findings are rather modest when viewed against the background of the massive array of extant insect species on Earth. Nevertheless, they underscore the significance of insect-derived substances for new drug discovery and development programs. Other examples besides those extensively addressed in this paper are the readily available and cost-effective anti-inflammatory glycosaminoglycans from the cricket Gryllus bimaculatus De Geer, 1773 (Orthoptera: Gryllidae) [247] and the powerful proteolytic enzyme mantis egg fibrolase (MEF) from the egg cases of the Chinese mantis Tenodera sinensis (Saussure, 1871) (Mantodea: Mantidae) [248]. Furthermore, products from the American cockroach Periplaneta americana Linnaeus, 1758 (Blattodea: Blattidae) [249], certain dipteran species [250], and the larvae of the dobsonfly Protohermes grandis Thunberg, 1781 (Megaloptera: Corydalidae) [251] displayed broad-spectrum in vitro antibiotic activity, including activity against methicillin-resistant Staphylococcus aureus. Notably, the insect antimicrobial peptide cecropin from the housefly Musca domestica Linnaeus, 1758 (Diptera: Muscidae) elicited interesting anticancer effects in a cell culture model [252]. And the development of bandages, gels, or lotions laced with purified debriding enzymes from the green bottle fly Lucilia sericata Meigen, 1826 (Calliphoridae) is anticipated to improve maggot debridement therapy [253].
However, drug discovery and development programs based on medicinal insects face a number of unprecedented obstacles. Firstly, the huge diversity of insect species makes it difficult to distinguish between species having potential therapeutic applicability and those with no apparent medicinal value. Indeed, with so many species altogether, it will take much time and many resources to separate the wheat from the chaff. Incidentally, this is probably an important reason for the failure of several bioprospecting endeavors such as the 1991-multimillion-dollar venture of the Costa Rican National Biodiversity Institute and the pharmaceutical giant Merck & Co. committed to the exploration of bioactive compounds from, among others, insects in the Costa Rican rainforests [254].
Secondly, insects are relatively small and, accordingly, produce only minute amounts of bioactive compounds. For instance, certain blister beetles have cantharidin loads of up to 35 μg per individual [26], and a honey bee holds 0.15 to 0.3 mg of venom in its venom sac, about half of it representing mellitin [255]. This necessitates the collection of vast numbers of animals to obtain sufficient material for mechanistic evaluations, animal pharmacological studies, and if warranted, clinical trials. Obviously, such an endeavor is highly problematic when the bioactive compound of interest is produced by species living solitatary or in small groups. As well, many natural products including those from insect have very complex structures, making total synthesis not always feasible or cost-effective [256], while culturing of the desired insects is in many cases not a valid option due to the lack of information about the living conditions and the feeding habits of most species [11,12].
Furthermore, climate change, particularly global warming phenomena caused by certain human activities, may destroy the habitats of many insect species, threatening their continued existence [257]. And although the medicinal use of insects has been well-documented in, among others, India, China, and Korea [258-260], the preservation of traditional knowledge about medicinal insects is in many cultures far from guaranteed.
Clearly, convertion of the therapeutic potential of medicinal insects into contemporary efficacious medicinal applications will require a collaborative effort of governments, holders of traditional knowledge, scientists, and pharmaceutical companies. Hopefully, such enterprises will eventually overcome the tremendous obstacles mentioned above, and lead to the full exploration of the valuable compounds represented by this largely unexplored source of bioactive substances and the development of structurally novel and mechanistically unique drugs to more effectively manage human diseases.
References
- Farnsworth NR (1984) The role of medicinal plants in drug development. Natural Products and Drug Development 8-98. Ballière, Tindall, and Cox, London, UK.
- Cox PA (1994) The ethnobotanical approach to drug discovery: strengths and limitations. Ciba Found Symp 185: 25-36. [Crossref]
- Mans DRA (2013) From forest to pharmacy. Acad J Med Plants 1: 101-110.
- Mans DRA (2016) Exploring the global animal biodiversity in the search for new drugs – marine invertebrates. J Transl Sci 2: 170-179.
- Breckenridge A (2006) William Withering's legacy--for the good of the patient. Clin Med (Lond) 6: 393-397. [Crossref]
- Bowman WC (2006) Neuromuscular block. Br J Pharmacol 147 Suppl 1: S277-286. [Crossref]
- Hadden DR (2005) Goat’s rue – French lilac – Italian fitch – Spanish sainfoin: Gallega officinalis and metformin: the Edinburgh connection. J R Coll Physicians Edinb 35: 258-260. [Crossref]
- Demain AL, Elander RP (1999) The beta-lactam antibiotics: past, present, and future. Antonie Van Leeuwenhoek 75: 5-19. [Crossref]
- McGivern JG (2007) Ziconotide: a review of its pharmacology and use in the treatment of pain. Neuropsychiatr Dis Treat 3: 69-85. [Crossref]
- Fayette J, Coquard IR, Alberti L, Ranchère D, Boyle H, et al. (2005) ET-743: a novel agent with activity in soft tissue sarcomas. Oncologist 10: 827-832. [Crossref]
- Johnson NF, Triplehorn CA (2005) Borror and DeLong’s introduction to the study of insects, (7th edtn). Cengage Learning, Boston (MA), USA.
- Gullan PJ, Cranston PS (2014) The insects: an outline of entomology (5th edtn). John Wiley and Sons, Chichester, West Sussex, UK.
- Regier JC, Shultz JW, Zwick A, Hussey A, Ball B, et al. (2010) Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences. Nature 463: 1079-1083.
- Misof B, Liu S, Meusemann K, Peters RS, Donath A, et al. (2014) Phylogenomics resolves the timing and pattern of insect evolution. Science 346: 763-767. [Crossref]
- Laurent P, Braekman J-C, Daloze D (2005) The chemistry of pheromones and other semiochemicals II.. Top Curr Chem 240: 167–229.
- Blum MS (1981) Chemical defenses of arthropods. Academic Press, New York, USA.
- Prestwich GD, Chen D (1981) Soldier defense secretions of Trinervitermes bettonianus (Isoptera, Nasutitermitinae): chemical variation in allopatric populations. J Chem Ecol 7: 147–157.
- Edwards JS (1961) The action and composition of the saliva of an assassin bug Platymeris rhadamanthus Gaerst (Hemiptera, Reduviidae). J Exp Biol 38: 61-77.
- Jones TH, Clark DA, Edwards AA, Davidson DW, Spande TF, et al. (2004) The chemistry of exploding ants, Camponotus spp. (cylindricus complex). J Chem Ecol 30: 1479-1492. [Crossref]
- Dean J, Aneshansley DJ, Edgerton HE, Eisner T (1990) Defensive spray of the bombardier beetle: a biological pulse jet. Science 248: 1219-1221. [Crossref]
- Kalaichelvan T, Verma KK (2000) Fecal cover for eggs of Indian cassidines (Coleoptera, Chrysomelidae). Ins Environ 6: 41-42.
- Whitman DW, Billen JPJ, Alsop D, Blum MS (1991) Anatomy, ultrastruicture, and functional morphology of the metathoracic tracheal defensive glands of the grasshopper Romalea guttata. Can J Zool 69: 2100-2108.
- Hojo MK, Pierce NE2, Tsuji K3 (2015) Lycaenid caterpillar secretions manipulate attendant ant behavior. Curr Biol 25: 2260-2264. [Crossref]
- Sahayaraj K, Borgio JF, Muthukumar S, Anandh GP (2006). Antibacterial activity of Rhynocoris marginatus (FAB.) and Catamirus brevipennis (Servile) (Hemiptera: Reduviidae) venoms against human pathogens. J Venom Anim Toxins incl Trop Dis 12: 487-496.
- Oršolić N (2012) Bee venom in cancer therapy. Cancer Metastasis Rev 31: 173-194. [Crossref]
- Ghoneim K (2014) Cantharidin as promising chemotherapeutic agent. J Sci 4: 272–292. [Crossref]
- Chakravorty J, Ghosh S, Meyer-Rochow VB (2011) Practices of entomophagy and entomotherapy by members of the Nyishi and Galo tribes, two ethnic groups of the state of Arunachal Pradesh (North-East India). J Ethnobiol Ethnomed 7: 5. [Crossref]
- Costa-Neto EM (2002) The use of insects in folk medicine in the state of Bahia, northeastern Brazil, with notes on insects reported elsewherein Brazilian folk medicine. Hum Ecol 30: 245-264.
- Ramos-Elorduy de Concini J, Pino Moreno JM (1988) The utilization of insects in the empirical medicine of ancient Mexicans. J Ethnobiol 8: 195-202.
- Bidau CJ (2014) Patterns in Orthoptera biodiversity. I. Adaptations in ecological and evolutionary contexts. J Insect Biodiv 2: 1-39.
- Whitman DW, Jones CG, Blum MS (1992) Defensive secretion production in lubber grasshoppers (Orthoptera: Romaleidae): influence of age, sex, diet, and discharge frequency. Ann Entomol Soc Am 85: 96–102.
- Whitman DW, Blum MS, Jones CG (1985) Chemical defense in Taeniopoda eques (Orthoptera: Acrididae): role of the metathoracic secretion. Ann Entomol Soc Am 78: 451–455.
- Whitman DW, Blum MS, Alsop DW (1990) Allomones: Chemicals for Defense. pp. 289–353. In: Evans DL, Schmidt JO, eds. Insect Defenses: Adaptive Mechanisms and Strategies of Prey and Predators. Albany, State University of New York, USA.
- Scherer CW, Capinera JL. Eastern lubber grasshopper, Romalea microptera (Beauvois) (= guttata (Houttuyn)) (Insecta: Orthoptera: Acrididae).
- Ingrassia L, Lefranc F, Mathieu V, Darro F, Kiss R (2008) Amaryllidaceae isocarbostyril alkaloids and their derivatives as promising antitumor agents. Transl Oncol 1: 1-13. [Crossref]
- Ji YB, Zheng J, Chen N, Song DX, Dong Y, et al. (2013) Isocarbostyril alkaloids and their derivatives as promising antitumor agents. Appl Mech Mat 411: 3150-3153.
- Hartwell JL (1967) Plants used against cancer. A survey. Lloydia 30: 379–436.
- Ceriotti G (1967) Narciclasine: an antimitotic substance from Narcissus bulbs. Nature 213: 595-596. [Crossref]
- Pettit GR, Pettit GR 3rd, Backhaus RA, Boyd MR, Meerow AW (1993) Antineoplastic agents, 256. Cell growth inhibitory isocarbostyrils from Hymenocallis. J Nat Prod 56: 1682-1687. [Crossref]
- Weniger B, Italiano L, Beck JP, Bastida J, Bergoñon S, et al. (1995) Cytotoxic activity of Amaryllidaceae alkaloids. Planta Med 61: 77-79. [Crossref]
- Pettit GR, Melody N (2005) Antineoplastic agents. 527. Synthesis of 7-deoxynarcistatin, 7-deoxy-trans-dihydronarcistatin, and trans-dihydronarcistatin 1(1). J Nat Prod 68: 207-211. [Crossref]
- Van Goietsenoven G, Mathieu V, Lefranc F, Kornienko A, Evidente A, et al. (2013) Narciclasine as well as other Amaryllidaceae isocarbostyrils are promising GTP-ase targeting agents against brain cancers. Med Res Rev 33: 439-455. [Crossref]
- Dumont P, Ingrassia L, Rouzeau S, Ribaucour F, Thomas S, et al. (2007) The Amaryllidaceae isocarbostyril narciclasine induces apoptosis by activation of the death receptor and/or mitochondrial pathways in cancer cells but not in normal fibroblasts. Neoplasia 9: 766–776. [Crossref]
- Kekre N, Griffin C, McNulty J, Pandey S (2005) Pancratistatin causes early activation of caspase-3 and the flipping of phosphatidyl serine followed by rapid apoptosis specifically in human lymphoma cells. Cancer Chemother Pharmacol 56: 29-38. [Crossref]
- McLachlan A, Kekre N, McNulty J, Pandey S (2005) Pancratistatin: a natural anti-cancer compound that targets mitochondria specifically in cancer cells to induce apoptosis. Apoptosis 10: 619-630. [Crossref]
- Griffin C, Sharda N, Sood D, Nair J, McNulty J, et al. (2007) Selective cytotoxicity of pancratistatin-related natural Amaryllidaceae alkaloids: evaluation of the activity of two new compounds. Cancer Cell Int 7:10. [Crossref]
- Griffin C, Hamm C, McNulty J, Pandey S (2010) Pancratistatin induces apoptosis in clinical leukemia samples with minimal effect on non-cancerous peripheral blood mononuclear cells. Cancer Cell Int 10: 6. [Crossref]
- Pettit GR, Meng Y, Herald DL, Knight JC, Day JF (2005) Antineoplastic agents 553. The Texas grasshopper. Brachystola magna. J Nat Prod 68: 1256-1258. [Crossref]
- Bright KL, Bernays EA, Moran VC (1994) Foraging patterns and dietary mixing in the field by the generalist grasshopper Brachystola magna (Orthoptera, Acrididae). J Insect Behavior 7: 779-793.
- Bordereau C, Robert A, van Tuyen V, Peppuy A (1997) Suicidal defensive behaviour by frontal gland dehiscence in Globitermes sulphureus Haviland soldiers (Isoptera). Insectes Soc 44: 289–297.
- Šobotník J, Bourguignon T, Hanus R, Weyda F, Roisin Y (2010). Structure and function of defensive glands in soldiers of Glossotermes oculatus (Isoptera: Serritermitidae). Biol J Linnean Soc 99: 839–848.
- Sobotník J, Jirosová A, Hanus R (2010) Chemical warfare in termites. J Insect Physiol 56: 1012-1021. [Crossref]
- Šobotník J, Bourguignon T, Hanus R, Demianová Z, Pytelková J, et al. (2012) Explosive backpacks in old termite workers. Science 337: 436. [Crossref]
- Coutinho HD, Lôbo KM, Bezerra DA, Lôbo I (2008) Peptides and proteins with antimicrobial activity. Indian J Pharmacol 40: 3-9. [Crossref]
- Srivastava SK, Babu N, Pandey H (2009) Traditional insect bioprospecting. As human food and medicine. Indian J Tradit Know 8: 485-494.
- Bulmer MS, Crozier RH (2006) Variation in positive selection in termite GNBPs and Relish. Mol Biol Evol 23: 317-326. [Crossref]
- Lamberty M, Zachary D, Lanot R, Bordereau C, Robert A, et al. (2001) Insect immunity, constitutive expression of a cysteine-rich antifungal and a linear antibacterial peptide in a termite insect. J Biol Chem 276: 4085-4092. [Crossref]
- Coutinho HDM, Vasconcellos A, Lima MA, Almeida-Filho GG, Alves RRN (2009) Termite usage associated with antibiotic therapy: enhancement of aminoglycoside antibiotic activity by natural products of Nausitermes corniger (Motschulsky 1855). BMC Complement Altern Med 9:35. [Crossref]
- Traniello JF, Rosengaus RB, Savoie K (2002) The development of immunity in a social insect: evidence for the group facilitation of disease resistance. Proc Natl Acad Sci USA 99: 6838-6842. [Crossref]
- Landon C, Meudal H, Boulanger N, Bulet P, Vovelle F (2006) Solution structures of stomoxyn and spinigerin, two insect antimicrobial peptides with an alpha-helical conformation. Biopolymers 81: 92-103. [Crossref]
- Coutinho HD1, Vasconcellos A, Freire-Pessôa HL, Gadelha CA, Gadelha TS, et al. (2010) Natural products from the termite Nasutitermes corniger lowers aminoglycoside minimum inhibitory concentrations. Pharmacogn Mag 6: 1-4. [Crossref]
- Chaves TP, Clementine ELC, Felimino DC, Alves RRN, Vasconcellos A, et al. (2015) Antibiotic resistance modulation by natural products obtained from Nasutitermis corniger Motschulsky, 1855 and its nest. Saudi J Biol Sci 22: 404-408. [Crossref]
- Sardar AH, Das S, Agnihorti S, Kumar M, Ghosh AK, et al. (2013) Spinigerin induces apoptotic like cell death in a caspase independent manner in Leishmania donovani. Exp Parasitol 135: 715-725. [Crossref]
- Kromp B (1999) Carabid beetles in sustainable agriculture: a review on pest control efficacy, cultivation impacts and enhancement. Agric Ecosyst Environ 74: 187-228.
- Carrel JE, McCairel MH, Slagle AJ, Doom JP, Brill J, et al. (1993) Cantharidin production in a blister beetle. Experientia 49: 171-174. [Crossref]
- Carrel JE, Doom JP, McCormick JP (1986) Identification of cantharidin in false blister beetles (Coleoptera, Oedemeridae) from Florida. J Chem Ecol 12: 741-747. [Crossref]
- McCluskey A, Walkom C, Bowyer MC, Ackland SP, Gardiner E et al. (2001) Cantharimides: a new class of modified cantharidin analogues inhibiting protein phosphatases 1 and 2A. Bioorg Med Chem Lett 11: 2941–2946. [Crossref]
- [Crossref] Nakatani T1, Konishi T, Miyahara K, Noda N (2004) Three novel cantharidin-related compounds from the Chinese blister beetle, Mylabris phalerata Pall. Chem Pharm Bull (Tokyo) 52: 807-809.
- Foster RW, Benhamou L, Porter MJ, Bucar DK, Hailes HC, et al. (2015) Irreversible endo-selective Diels-Alder reactions of substituted alkoxyfurans: a general synthesis of endo-cantharimides. Chemistry 21: 6107-6014. [Crossref]
- Lin P-Y, Shi S-J, Hsu F-L, Chen C-F (1998) New cantharidinimides from cantharidin and 2-arylethylamines: efficient synthesis under high pressure. J Chinese Chem Soc 45: 323–326.
- Nikbakhtzadeh MR, Ebramihi B (2007) Detection of cantharidin-related compounds in Mylabris impressa (Coleoptera: Meloidae). J Venom Anim Toxins Incl Trop Dis 13: 686–693.
- Chen YJ, Tsai YM, Kuo CD, Ku KL, Shie HS, et al. (2009) Norcantharidin is a small-molecule synthetic compound with anti-angiogenesis effect. Life Sci 85: 642-651. [Crossref]
- Moed L, Shwayder TA, Chang MW (2001) Cantharidin revisited: a blistering defense of an ancient medicine. Arch Dermatol 137: 1357-1360. [Crossref]
- Wang GS (1989) Medical uses of mylabris in ancient China and recent studies. J Ethnopharmacol 26: 147-162. [Crossref]
- Mathes EF, Frieden IJ (2010) Treatment of molluscum contagiosum with cantharidin: a practical approach. Pediatr Ann 39: 124-130. [Crossref]
- Torbeck R, Pan M, de Moll E, Levitt J (2014) Cantharidin: a comprehensive review of the clinical literature. Dermatol Online J 20: 3. [Crossref]
- Rauh R, Kahl S, Boechzelt H, Bauer R, Kaina B, et al. (2007) Molecular biology of cantharidin in cancer cells. Chin Med 2: 8. [Crossref]
- Liu D, Chen Z (2009) The effects of cantharidin and cantharidin derivates on tumour cells. Anticancer Agents Med Chem 9: 392-396. [Crossref]
2021 Copyright OAT. All rights reserv
- Deng LP, Dong J, Cai H, Wang W (2013) Cantharidin as an antitumor agent: a retrospective review. Curr Med Chem 20: 159-166. [Crossref]
- Hong CY, Huang SC, Lin SK, Lee JJ, Chueh LL, et al. (2000) Norcantharidin-induced post-G(2)/M apoptosis is dependent on wild-type p53 gene. Biochem Biophys Res Commun 276: 278-285. [Crossref]
- Kuo JH, Chu YL, Yang JS, Lin JP, Lai KC, et al. (2010) Cantharidin induces apoptosis in human bladder cancer TSGH 8301 cells through mitochondria-dependent signal pathways. Int J Oncol 37: 1243-1250. [Crossref]
- Shen B, He P-J, Shao C-L (2013) Norcantharidin-induced DU145 cell apoptosis through ROS-mediated mitochondrial dysfunction and energy depletion. PLoS ONE 8: e84610. [Crossref]
- Tian X, Zeng G, Li X, Wu Z, Wang L (2015) Cantharidin inhibits cell proliferation and promotes apoptosis in tongue squamous cell carcinoma through suppression of miR-214 and regulation of p53 and Bcl-2/Bax. Oncol Rep 33: 3061-3068. [Crossref]
- Li W, Xie L, Chen Z, Zhu Y, Sun Y, et al. (2010) Cantharidin, a potent and selective PP2A inhibitor, induces an oxidative stress-independent growth inhibition of pancreatic cancer cells through G2/M cell-cycle arrest and apoptosis. Cancer Sci 101: 1226–1233. [Crossref]
- Yu T, Hou F, Liu M, Zhou L, Li D, et al. (2012) Norcantharidin anti-angiogenesis activity possibly through an endothelial cell pathway in human colorectal cancer. Asian Pac J Cancer Prev 13: 499-503. [Crossref]
- Zhang L, Ji Q, Liu X, Chen X, Chen Z, et al. (2013) Norcantharidin inhibits tumor angiogenesis via blocking VEGFR2/MEK/ERK signaling pathways. Cancer Sci 104: 604-610. [Crossref]
- Fan Y-Z, Fu J-Y, Zhao Z-M, Chen C-Q (2007) Inhibitory effect of norcantharidin on the growth of human gallbladder carcinoma GBC-SD cells in vitro. Hepatobiliary Pancreat Dis Int 6: 72–80. [Crossref]
- Wang H, Sun W, Zhang WZ, Ge CY, Zhang JT, et al. (2014) Inhibition of tumor vasculogenic mimicry and prolongation of host survival in highly aggressive gallbladder cancers by norcantharidin via blocking the ephrin type a receptor 2/focal adhesion kinase/paxillin signaling pathway. PLoS One 9: e96982. [Crossref]
- Li CC, Yu FS, Fan MJ, Chen YY, et al. (2016) Anticancer effects of cantharidin in A431 human skin cancer (Epidermoid carcinoma) cells in vitro and in vivo. Environ Toxicol. [Crossref]
- Zhang JT, Sun W, Zhang WZ, Ge CY, Liu ZY, et al. (2014) Norcantharidin inhibits tumor growth and vasculogenic mimicry of human gallbladder carcinomas by suppression of the PI3-K/MMPs/Ln-5y2 signaling pathway. BMC Cancer 14: 193.
- Chen Y-J, Kuo C-D, Tsai Y-M, Yu C-C, Wang G-S, et al. (2008) Norcantharidin induces anoikis through Jun-N terminal kinase activation in CT26 colorectal cancer cells. Anticancer Drugs 19: 55-64. [Crossref]
- Peng C, Li Z, Niu Z, Niu W, Xu Z, et al. (2016) Norcantharidin suppresses colon cancer cell epithelial-mesenchymal transition by inhibiting the αvβ6-ERK-Ets1signaling pathway. Sci Rep 6: 20500. [Crossref]
- Li YM, Casida JE (1992) Cantharidin-binding protein: identification as protein phosphatase 2A. Proc Natl Acad Sci USA 89: 11867-11870. [Crossref]
- Honkanen RE (1993) Cantharidin, another natural toxin that inhibits the activity of serine/threonine protein phosphatases types 1 and 2A. FEBS Lett 330: 283-286. [Crossref]
- Ciccone M, Calin GA, Perrotti D (2015) From the Biology of PP2A to the PADs for therapy of hematologic malignancies. Front Oncol 5: 21. [Crossref]
- Cui DL, Yao XX (2008) Progress in the treatment of hepatitis B and digestive tumors with cantharidin and its analogues. World Chinese J Digestol 13: 5-13.
- Zhan Y-P, Huang X-E, Cao J, Lu Y-Y, Wu X-Y, et al. (2012) Clinical study on safety and efficacy of Qinin® (cantharidin sodium) injection combined with chemotherapy in treating patients with gastric cancer. Asian Pac J Cancer Prev 13: 4773-4776. [Crossref]
- Dang YJ, Zhu CY (2013) Oral bioavailability of cantharidin-loaded solid lipid nanoparticles. Chin Med 8: 1. [Crossref]
- Lin LH, Huang HS, Lin CC, Lee LW, Lin PY (2004) Effects of cantharidinimides on human carcinoma cells. Chem Pharm Bull (Tokyo) 52: 855-857. [Crossref]
- Yang H, Guo W, Xu B, Li M, Cui J (2007) Anticancer activity and mechanisms of norcantharidin-Nd3II on hepatoma. Anticancer Drugs 18: 1133-1137. [Crossref]
- Yi SN, Wass J, Vincent P, Iland H (1991) Inhibitory effect of norcantharidin on K562 human myeloid leukemia cells in vitro. Leuk Res 15: 883-886. [Crossref]
- Kok S-H, Cheng S-J, Hong C-Y, Lee J-J, Lin S-K, et al. (2005) Norcantharidin-induced apoptosis in oral cancer cells is associated with an increase of proapoptotic to antiapoptotic protein ratio. Cancer Lett 217: 43–52. [Crossref]
- Chen YJ, Kuo CD, Chen SH, Chen WJ, Huang WC, et al. (2012) Small-molecule synthetic compound norcantharidin reverses multi-drug resistance by regulating sonic hedgehog signaling in human breast cancer cells. PLoS One 75:e37006. [Crossref]
- Dorn DC, Kou CA, Png KJ, Moore MA (2009) The effect of cantharidins on leukemic stem cells. Int J Cancer 124: 2186-2199. [Crossref]
- Zhang J, Tang Y, Li S, Liao, Guo X (2010) Targeting of the B-lineage leukemia stem cells and their progeny with norcantharidin encapsulated liposomes modified with a novel CD19 monoclonal antibody 2E8 in vitro. J Drug Target 18: 675–687. [Crossref]
- Liao HF, Chen YJ, Chou CH, Wang FW, Kuo CD (2011) Norcantharidin induces cell cycle arrest and inhibits progression of human leukemic Jurkat T cells through mitogen-activated protein kinase-mediated regulation of interleukin-2 production. Toxicol In Vitro 25: 206-212. [Crossref]
- Yeh CB, Hsieh MJ, Hsieh YH, Chien MH, Chiou HL, et al. (2012) Antimetastatic effects of norcantharidin on hepatocellular carcinoma by transcriptional inhibition of MMP-9 through modulation of NF-kB activity. PLoS One 7: e31055. [Crossref]
- Huang Y, Liu Q, Liu K, Yagasaki K, Zhang G (2009) Suppression of growth of highly-metastatic human breast cancer cells by norcantharidin and its mechanisms of action. Cytotechnology 59: 201-208. [Crossref]
- Liu D, Chen Z (2009) The effects of cantharidin and cantharidin derivates on tumour cells. Anticancer Agents Med Chem 9: 392-396. [Crossref]
- Munstermann LE, Conn JE (1997) Systematics of mosquito disease vectors (Diptera, Culicidae): impact of molecular biology and cladistic analysis. Annu Rev Entomol 42: 351-369. [Crossref]
- Kopecký J, Kuthejlová M (1998) Suppressive effect of Ixodes ricinus salivary gland extract on mechanisms of natural immunity in vitro. Parasite Immunol 20: 169–174. [Crossref]
- Arcá B, Lombardo F, de Lara Capurro M, della Torre A, Dimopoulos G, et al. (1999) Trapping cDNAs encoding secreted proteins from the salivary glands of the malaria vector Anopheles gambiae. Proc Natl Acad Sci USA 96: 1516–1521. [Crossref]
- Charlab R, Valenzuela JG, Rowton ED, Ribeiro JM (1999) Toward an understanding of the biochemical and pharmacological complexity of the saliva of a hematophagous sand fly Lutzomyia longipalpis. Proc Natl Acad Sci USA 96: 15155-15160. [Crossref]
- Ribeiro JM, Francischetti IM (2003) Role of arthropod saliva in blood feeding: sialome and post-sialome perspectives. Annu Rev Entomol 48: 73-88. [Crossref]
- Ma D, Wang Y, Yang H, Wu J, An S, et al. (2009) Anti-thrombosis repertoire of blood-feeding horsefly salivary glands. Mol Cell Proteomics 8: 2071-2079. [Crossref]
- Yang X, Hu K, Yan G, Zhang Y, Pei Y (2000) Fibronogenolytic componments in Tabanid'an ingredient in traditional Chinese medicine and their properties (Chinese). J Southwest Agricultural University 22: 173–176.
- Adams RL, Bird RJ (2009) Coagulation cascade and therapeutics update: relevance to nephrology. Part 1: Overview of coagulation, thrombophilias and history of anticoagulants. Nephrology (Carlton) 14: 462-470. [Crossref]
- Crawley JT, Lane DA (2008) The haemostatic role of tissue factor pathway inhibitor. Arterioscler Thromb Vasc Biol 28: 233-242. [Crossref]
- Corral-Rodríguez MA, Macedo-Ribeiro S, Barbosa Pereira PJ, Fuentes-Prior P (2009) Tick-derived Kunitz-type inhibitors as antihemostatic factors. Insect Biochem Mol Biol 39: 579-595. [Crossref]
- Francischetti IM, Mather TN, Ribeiro JM (2004) Penthalaris, a novel recombinant five-Kunitz tissue factor pathway inhibitor (TFPI) from the salivary gland of the tick vector of Lyme disease, Ixodes scapularis. Thromb Haemost 91: 886-898. [Crossref]
- Lane RS, Piesman J, Burgdorfer W (1991) Lyme borreliosis: relation of its causative agent to its vectors and hosts in North America and Europe. Annu Rev Entomol 36: 587-609. [Crossref]
- Jacobs JW, Cupp EW, Sardana M, Friedman PA (1990) Isolation and characterization of a coagulation factor Xa inhibitor from black fly salivary glands. Thromb Haemost 64: 235-238. [Crossref]
- Abebe M, Cupp MS, Ramberg FB, Cupp EW (1994) Anticoagulant activity in salivary gland extracts of black flies (Diptera: Simuliidae). J Med Entomol 31: 908-911. [Crossref]
- Abebe M, Cupp MS, Champagne D, Cupp EW (1995) Simulidin: a black fly (Simulium vittatum) salivary gland protein with anti-thrombin activity. J Insect Physiol 41: 1001–1006.
- Abebe M, Ribeiro JMC, Cupp MS, Cupp EW (1996) Novel anticoagulant from salivary glands of Simulium vittatum (Diptera: Simuliidae) inhibits activity of coagulation factor V. J Med Entomol 33: 173–176.
- Diemert DJ (2011) Tissue nematode infections. Goldman’s Cecil Medicine 193-200.
- Andersen JF, Pham VM, Meng Z, Champagne DE, Ribeiro JM (2009) Insight into the sialome of the black fly, Simulium vittatum. J Proteome Res 8: 1474-1488. [Crossref]
- Tsujimoto H, Kotsyfakis M, Francischetti IMB, Eum JH, Strand MR, et al. (2012) Simukunin from the salivary glands of the black fly Simulium vittatum inhibits enzymes that regulate clotting and inflammatory responses. PLoS ONE 7: e29964. [Crossref]
- Merelo-Lobo AR, McCall PJ, Perez MA, Spiers AA, Mzilahowa T, et al. (2003) Identification of the vectors of lymphatic filariasis in the Lower Shire Valley, southern Malawi. Trans R Soc Trop Med Hyg 97: 299-301. [Crossref]
- Ma D, Xu X, An S, Liu H, Yang X, et al. (2011) A novel family of RGD-containing disintegrin (Tablysin-15) from the salivary gland of the horsefly Tabanus yao targets integrins aIIbß3 and aVß3 and inhibits platelet aggregation and angiogenesis. Thromb Haemost 105: 1032–1045. [Crossref]
- Valenzuela JG, Francischetti IM, Ribeiro JM (1999) Purification, cloning, and synthesis of a novel salivary anti-thrombin from the mosquito Anopheles albimanus. Biochemistry 38: 11209-11215. [Crossref]
- Ronca R, Kotsyfakis M, Lombardo F, Rizzo C, Currà C, et al. (2012) The Anopheles gambiae cE5, a tight- and fast-binding thrombin inhibitor with post-transcriptionally regulated salivary-restricted expression. Insect Biochem Mol Biol 42: 610–620. [Crossref]
- Andersen JF, Gudderra NP, Francischetti IM, Ribeiro JM (2005) The role of salivary lipocalins in blood feeding by Rhodnius prolixus. Arch Insect Biochem Physiol 58: 97-105. [Crossref]
- Isawa H, Yuda M, Yoneda K, Chinzei Y (2000) The insect salivary protein, prolixin-S, inhibits factor IXa generation and Xase complex formation in the blood coagulation pathway. J Biol Chem 275: 6636–6641. [Crossref]
- Gudderra NP, Ribeiro JM, Andersen JF (2005) Structural determinants of factor IX(a) binding in nitrophorin 2, a lipocalin inhibitor of the intrinsic coagulation pathway. J Biol Chem 280: 25022-25028. [Crossref]
- Kaneko Y, Shojo H, Yuda M, Chinzei Y (1999) Effects of recombinant nitrophorin-2 nitric oxide complex on vascular smooth muscle. Biosci Biotechnol Biochem 63: 1488-1490. [Crossref]
- Montfort WR, Weichsel A, Andersen JF (2000) Nitrophorins and related antihemostatic lipocalins from Rhodnius prolixus and other blood-sucking arthropods. Biochim Biophys Acta 1482: 110–118. [Crossref]
- Carneiro-Lobo TC, Konig S, Machado DE, Nasciutti LE, Forni MF, et al. (2009) Ixolaris, a tissue factor inhibitor, blocks primary tumor growth and angiogenesis in a glioblastoma model. J Thromb Haemost 7: 1855-1864. [Crossref]
- Carneiro-Lobo TC, Schaffner F, Disse J, Ostergaard H, Francischetti IMB, et al. (2012) The tick-derived inhibitor Ixolaris prevents tissue factor signaling on tumor cells. Thromb Haemost 10: 1849–1858. [Crossref]
- Da Silva de Oliveira A, Lima LG, Mariano-Oliveira A, Machado DE, Nasciutti LE, et al. (2012) Inhibition of tissue factor by ixolaris reduces primary tumor growth and experimental metastasis in a murine model of melanoma. Thromb Res 130: e163–e170. [Crossref]
- Hughes DG, Bennett, PM (1991) Captive breeding and the conservation of invertebrates. Int Zoo Year Book 30: 45-51.
- Pierce NE (1995) Predatory and parasitic Lepidoptera: carnivores living on plants. J Lepidopterist's Soc 49: 412–453.
- Terren M, Mignon J, Declerck C, Jijakli H, Savery S, et al. (2012) Principal disease and insect pests of Jatropha curcas L. in the lower valley of the Senegal River. Tropicultura 30: 222-229.
- Koyama K, Wakabayashi K, Masutani M, Koiwai K, Watanabe M, et al. (1996) Presence in Pieris rapae of cytotoxic activity against human carcinoma cells. Jpn J Cancer Res 87: 1259-1262. [Crossref]
- Watanabe M1, Kono T, Koyama K, Sugimura T, Wakabayashi K (1998) Purification of pierisin, an inducer of apoptosis in human gastric carcinoma cells, from cabbage butterfly, Pieris rapae. Jpn J Cancer Res 89: 556-561. [Crossref]
- Watanabe M, Kono T, Matsushima-Hibiya Y, Kanazawa T, Nishisaka N, et al. (1999) Molecular cloning of an apoptosis inducing protein, pierisin, from cabbage butterfly: possible involvement of ADP-ribosylation in its activity. Proc Natl Acad Sci USA 96: 10608-10613. [Crossref]
- Matsushima-Hibiya Y1, Watanabe M, Kono T, Kanazawa T, Koyama K, et al. (2000) Purification and cloning of pierisin-2, an apoptosis-inducing protein from the cabbage butterfly, Pieris brassicae. Eur J Biochem 267: 5742-5750. [Crossref]
- Takahashi-Nakaguchi A, Matsumoto Y, Yamamoto M, Iwabuchi K, Totsuka Y, et al. (2013) Demonstration of cytotoxicity against wasps by pierisin-1: a possible defense factor in the cabbage white butterfly. PLoS One 8: e60539. [Crossref]
- Kono T, Watanabe M, Koyama K, Kishimoto T, Fukushima S, et al. (1999) Cytotoxic activity of pierisin, from the cabbage butterfly, Pieris rapae, in various human cancer cell lines. Cancer Lett 137: 75-81. [Crossref]
- Kanazawa T, Watanabe M, Matsushima-Hibiya Y, Kono T, Tanaka N, et al. (2001) Distinct roles for the N- and C-terminal regions in the cytotoxicity of pierisin-1, a putative ADP-ribosylating toxin from cabbage butterfly, against mammalian cells. Proc Natl Acad Sci USA 98: 2226–2231. [Crossref]
- Takamura-Enya T, Watanabe M, Koyama K, Sugimura T, Wakabayashi K (2004) Mono(ADP-ribosyl)ation of the N2 amino groups of guanine residues in DNA by pierisin-2, from the cabbage butterfly, Pieris brassicae. Biochem Biophys Res Commun 323: 579-582. [Crossref]
- Subbarayan S, Marimuthu SK, Nachimuthu SK, Zhang W, Subramanian S (2016) Characterization and cytotoxic activity of apoptosis-inducing pierisin-5 protein from white cabbage butterfly. Int J Biol Macromol 87: 16-27. [Crossref]
- Kanazawa T, Kono T, Watanabe M, Matsushima-Hibiya Y, Nakano T, et al. (2002) Bcl-2 blocks apoptosis caused by pierisin-1, a guanine-specific ADP-ribosylating toxin from the cabbage butterfly. Biochem Biophys Res Commun 296: 20-25.
- Watanabe M, Enomoto S, Takamura-Enya T, Nakano T, Koyama K, et al. (2004) Enzymatic properties of pierisin-1 and its N-terminal domain, a guanine-specific ADP-ribosyltransferase from the cabbage butterfly. J Biochem 135: 471-477. [Crossref]
- Watanabe M, Nakano T, Shiotani B, Matsushima-Hibiya Y, Kiuchi M, et al. (2004) Developmental stage-specific expression and tissue distribution of pierisin-1, a guanine-specific ADP-ribosylating toxin, in Pieris rapae. Comp Biochem Physiol A Mol Integr Physiol 139: 125-131. [Crossref]
- Spillner E, Blank S, Jakob T (2014) Hymenoptera allergens: from venom to "venome". Front Immunol 5: 77. [Crossref]
- Anderson KE, Eckholm B, Mott BM, Sheehan TH, Hoffman GD (2011) An emerging paradigm of colony health: microbial balance of the honey bee and hive (Apis mellifera). Insectes Soc 19: 431–444.
- Lubke LL, Garon CF (1997) The antimicrobial agent melittin exhibits powerful inhibitory effects on the Lyme disease spirochete. Clin Infect Dis 25 (Suppl 1): S48–S51. [Crossref]
- Klotz SA, Gaur NK, Rauceo J, Lake DF, Park Y, et al (2004) Inhibition of adherence and killing of Candida albicans with a 23-Mer peptide (Fn/23) with dual antifungal properties. Antimicrob Agents Chemother 48: 4337-4341. [Crossref]
- Lazarev VN, Shkarupeta MM, Titova GA, Kostrjukova ES, Akopian TA, et al. (2005) Effect of induced expression of an antimicrobial peptide melittin on Chlamydia trachomatis and Mycoplasma hominis infections in vivo. Biochem Biophys Res Commun 338: 946-950. [Crossref]
- De Lima PR, Brochetto-Braga MR (2003) Hymenoptera venom review focusing on Apis mellifera. J Venom Anim Toxins incl Trop Dis 9: 149-162.
- Raghuraman H1, Chattopadhyay A (2007) Melittin: a membrane-active peptide with diverse functions. Biosci Rep 27: 189-223. [Crossref]
- Monteiro MC1, Romão PR, Soares AM (2009) Pharmacological perspectives of wasp venom. Protein Pept Lett 16: 944-952. [Crossref]
- Shen LR, Ren YC (1999) Advances on exploration and utilization of the edible and medical ants in China. Entomol Knowl 36: 251-254.
- Kou J, Ni Y, Li N, Wang J, Liu L, et al. (2005) Analgesic and anti-inflammatory activities of total extract and individual fractions of Chinese medicinal ants Polyrhachis lamellidens. Biol Pharm Bull 28: 176-180.
- Feng Y, Zhao M, He Z, Chen Z, Sun L (2009) Research and utilization of medicinal insects in China. Entomol Res 39: 313-316.
- Hoffman DR (2006) Hymenoptera venom allergens. Clin Rev Allergy Immunol 30: 109-128. [Crossref]
- Munstedt J, Hackethal A, Schmidt K (2005) Bee venom therapy, bee venom acupuncture of apiculture: what is the evidence behind the various health claims? Am Bee J 145: 665–668.
- Oršolić N (2012) Bee venom in cancer therapy. Cancer Metastasis Rev 31: 173-194. [Crossref]
- Han SM, Lee KG, Pak SC (2013) Effects of cosmetics containing purified honeybee (Apis mellifera L.) venom on acne vulgaris. J Integr Med 11: 320-326. [Crossref]
- Cho SY, Shim SR, Rhee HY, Park HJ, Jung WS, et al. (2012) Effectiveness of acupuncture and bee venom acupuncture in idiopathic Parkinson's disease. Parkinsonism Relat Disord 18: 948-952. [Crossref]
- Shimpi R, Chaudhari P, Deshmukh R, Devare S, Bagad Y, et al. (2016) A review: pharmacotherapeutics of bee venom. World J Pharm Pharmaceut Sci 5: 656-667.
- Lee MS, Pittler MH, Shin BC, Kong JC, Ernst E (2008) Bee venom acupuncture for musculoskeletal pain: a review. J Pain 9: 289-297. [Crossref]
- Lee JA, Son MJ, Choi J, Jun JH, Kim JI, et al. (2014) Bee venom acupuncture for rheumatoid arthritis: a systematic review of randomised clinical trials. BMJ Open 4: e006140. [Crossref]
- Habermann E (1972) Bee and wasp venoms. Science 177: 314-322. [Crossref]
- Bernheimer AW, Rudy B (1986) Interactions between membranes and cytolytic peptides. Biochim Biophys Acta 864: 123-141. [Crossref]
- Dempsey CE (1990) The actions of melittin on membranes. Biochim Biophys Acta 1031: 143-161. [Crossref]
- Saberwal G, Nagaraj R (1994) Cell-lytic and antibacterial peptides that act by perturbing the barrier function of membranes: facets of their conformational features, structure–function correlation and membrane-perturbing abilities. Biochim Biophys Acta 1197: 109–131.
- Damianoglou A, Rodger A, Pridmore C, Dafforn TR, Mosely JA, et al. (2010) The synergistic action of melittin and phospholipase A2 with lipid membranes: development of linear dichroism for membrane-insertion kinetics. Protein Pept Lett 17: 1351–1362. [Crossref]
- Blondelle SE, Houghten RA (1991) Hemolytic and antimicrobial activities of the twenty-four individual omission analogues of melittin. Biochemistry 30: 4671-4678. [Crossref]
- Merrifield RB, Juvvadi P, Andreu D, Ubach J, Boman A, et al. (1995) Retro and retroenantio analogs of cecropin-melittin hybrids. Proc Natl Acad Sci USA 92: 3449-3453. [Crossref]
- Wachinger M, Saermark T, Erfle V (1992) Influence of amphipathic peptides on the HIV-1 production in persistently infected T lymphoma cells. FEBS Lett 309: 235-241. [Crossref]
- Baghian A, Jaynes J, Enright F, Kousoulas KG (1997) An amphipathic alpha-helical synthetic peptide analogue of melittin inhibits herpes simplex virus-1 (HSV-1)-induced cell fusion and virus spread. Peptides 18: 177–183. [Crossref]
- Song CC, Lu X, Cheng BB, DU J, Li B, et al. (2007) [Effects of melittin on growth and angiogenesis of human hepatocellular carcinoma BEL-7402 cell xenografts in nude mice]. Ai Zheng 26: 1315-1322. [Crossref]
- Gajski G, Garaj-Vrhovac V (2013) Melittin: a lytic peptide with anticancer properties. Environ Toxicol Pharmacol 36: 697-705. [Crossref]
- Liu S, Yu M, He Y, Xiao L, Wang F, et al. (2008) Melittin prevents liver cancer cell metastasis through inhibition of the Rac1-dependent pathway. Hepatology 47: 1964-1973. [Crossref]
- Huh JE, Kang JW, Nam D, Baek YH, Choi DY, et al. (2012) Melittin suppresses VEGF-A-induced tumor growth by blocking VEGFR-2 and the COX-2-mediated MAPK signaling pathway. J Nat Prod 75: 1922-1929. [Crossref]
- Shin JM, Jeong YJ, Cho HJ, Park KK, Chung IK, et al. (2013) Melittin suppresses HIF-1alpha/VEGF expression through inhibition of ERK and mTOR/p70S6K pathway in human cervical carcinoma cells. PLoS One 8: e69380. [Crossref]
- Jo M, Park MH, Kollipara PS, An BJ, Song HS, et al. (2012) Anti-cancer effect of bee venom toxin and melittin in ovarian cancer cells through induction of death receptors and inhibition of JAK2/STAT3 pathway. Toxicol Appl Pharmacol 258: 72-81. [Crossref]
- Yang X, Zhu H, Ge Y, Liu J, Cai J, et al. (2014) Melittin enhances radiosensitivity of hypoxic head and neck squamous cell carcinoma by suppressing HIF-1α. Tumour Biol 35: 10443-10448. [Crossref]
- Jin H, Li C, Li D, Cai M, Li Z, et al. (2013) Construction and characterization of a CTLA-4-targeted scFv-melittin fusion protein as a potential immunosuppressive agent for organ transplant. Cell Biochem Biophys 67: 1067–1074. [Crossref]
- Saini SS, Peterson JW, Chopra AK (1997) Melittin binds to secretory phospholipase A2 and inhibits its enzymatic activity. Biochem Biophys Res Commun 238: 436-442. [Crossref]
- Gabay C, Hasler P, Kyburz D, So A, Villiger P, et al. (2014) Biological agents in monotherapy for the treatment of rheumatoid arthritis. Swiss Med Wkly 144: w13950. [Crossref]
- Park HJ, Son DJ, Lee CW, Choi MS, Lee US, et al. (2007) Melittin inhibits inflammatory target gene expression and mediator generation via interaction with IkappaB kinase. Biochem Pharmacol 73: 237–247.
- Kim SJ, Park JH, Kim KH, Lee WR, Kim KS, et al. (2011) Melittin inhibits atherosclerosis in LPS/high-fat treated mice through atheroprotective actions. J Atheroscler Thromb 18: 1117-1126. [Crossref]
- Moreno M, Giralt E (2015) Three valuable peptides from bee and wasp venoms for therapeutic and biotechnological use: melittin, apamin and mastoparan. Toxins (Basel) 7: 1126-1150. [Crossref]
- Cerovský V, Budesínský M, Hovorka O, Cvacka J, Voburka Z, et al. (2009) Lasioglossins: three novel antimicrobial peptides from the venom of the eusocial bee Lasioglossum laticeps (Hymenoptera: Halictidae). Chembiochem 10: 2089-2099. [Crossref]
- Bandyopadhyay S, Lee M, Sivaraman J, Chatterjee C (2013) Model membrane interaction and DNA-binding of antimicrobial peptide Lasioglossin II derived from bee venom. Biochem Biophys Res Commun 430: 1-6. [Crossref]
- Slaninová J, Putnová H, Borovicková L, Šácha P, Cerovský V, et al. (2011) The antifungal effect of peptides from hymenoptera venom and their analogs. Cent Eur J Biol 6: 150–159.
- Hirai Y, Yasuhara T, Yoshida H, Nakajima T, Fujino M, et al. (1979) A new mast cell degranulating peptide "mastoparan" in the venom of Vespula lewisii. Chem Pharm Bull (Tokyo) 27: 1942-1944. [Crossref]
- Nakajima T (1984) Biochemistry of vespid venoms. In: Tu AT (ed). Handbook of Natural Toxins 109–133. Marcel Dekker, New York, USA.
- Nakajima T, Uzu S, Wakamatsu K, Saito K, Miyazawa T, et al. (1986) Amphiphilic peptides in wasp venom. Biopolymers 25 Suppl: S115-121. [Crossref]
- Higashijima T, Uzu S, Nakajima T, Ross EM (1988) Mastoparan, a peptide toxin from wasp venom, mimics receptors by activating GTP-binding regulatory proteins (G proteins). J Biol Chem 263: 6491-6494. [Crossref]
- Wakamatsu K, Higashijima T, Fujino M, Nakajima T, Miyazawa T, et al. (1983) Transferred NOE analyses of conformations of peptides as bound to membrane bilayer of phospholipid; mastoparan-X. FEBS Lett 162: 123–126.
- Katsu T, Kuroko M, Morikawa T, Sanchika K, Yamanaka H, et al. (1990) Interaction of wasp venom mastoparan with biomembranes. Biochim Biophys Acta 1027: 185-190. [Crossref]
- Nakahata N, Abe MT, Matsuoka I, Nakanishi H (1990) Mastoparan inhibits phosphoinositide hydrolysis via pertussis toxin-insensitive G-protein in human astrocytoma cells. FEBS Lett 260: 91–94. [Crossref]
- Ozaki Y, Matsumoto Y, Yatomi Y, Higashihara M, Kariya T, et al. (1990) Mastoparan, a wasp venom, activates platelets via pertussis toxin-sensitive GTP-binding proteins. Biochem Biophys Res Commun 170: 779–785. [Crossref]
- Weingarten R, Ransnäs L, Mueller H, Sklar LA, Bokoch GM (1990) Mastoparan interacts with the carboxyl terminus of the alpha subunit of Gi. J Biol Chem 265: 11044-11049. [Crossref]
- Lin SZ, Yan GM, Koch KE, Paul SM, Irwin RP (1997) Mastoparan-induced apoptosis of cultured cerebellar granule neurons is initiated by calcium release from intracellular stores. Brain Res 771: 184-195. [Crossref]
- Rocha T, de Souza BM, Palma MS, da Cruz-Höfling MA (2007) Myotoxic effects of mastoparan from Polybia paulista (Hymenoptera, Epiponini) wasp venom in mice skeletal muscle. Toxicon 50: 589-599. [Crossref]
- Rocha T, de Barros LL, Fontana K, de Souza BM, Palma MS, et al. (2010) Inflammation and apoptosis induced by mastoparan Polybia-MPII on skeletal muscle. Toxicon 55: 1213-1221. [Crossref]
- De Azevedo RA, Figueiredo CR, Ferreira AK, Matsuo AL, Massaoka MH, et al. (2015) Mastoparan induces apoptosis in B16F10-Nex2 melanoma cells via the intrinsic mitochondrial pathway and displays antitumor activity in vivo. Peptides 68: 113-119. [Crossref]
- Avram S, Buiu C, Borcan F, Milac AL (2012) More effective antimicrobial mastoparan derivatives, generated by 3D-QSAR-Almond and computational mutagenesis. Mol Biosyst 8: 587–594. [Crossref]
- Lin CH, Hou RF, Shyu CL, Shia WY, Lin CF, et al. (2012) In vitro activity of mastoparan-AF alone and in combination with clinically used antibiotics against multiple-antibiotic-resistant Escherichia coli isolates from animals. Peptides 36: 114–120. [Crossref]
- Sample CJ, Hudak KE, Barefoot BE, Koci MD, Wanyonyi MS, et al. (2013) A mastoparan-derived peptide has broad-spectrum antiviral activity against enveloped viruses. Peptides 48: 96–105. [Crossref]
- Henriksen JR, Etzerodt T, Gjetting T, Andresen TL (2014) Side chain hydrophobicity modulates therapeutic activity and membrane selectivity of antimicrobial peptide mastoparan-X. PLoS One 9: e91007. [Crossref]
- Danilenko M, Worland P, Carlson B, Sausville EA, Sharoni Y (1993) Selective effects of mastoparan analogs: separation of G-protein-directed and membrane-perturbing activities. Biochem Biophys Res Commun 196: 1296–1302. [Crossref]
- Wu TM, Li ML (1999) The cytolytic action of all-D mastoparan M on tumor cell lines. Int J Tissue React 21: 35-42. [Crossref]
- Jones S, Martel C, Belzacq-Casagrande AS, Brenner C, Howl J (2008) Mitoparan and target-selective chimeric analogues: membrane translocation and intracellular redistribution induces mitochondrial apoptosis. Biochim Biophys Acta 1783: 849–863. [Crossref]
- De Azevedo RA, Figueiredo CR, Ferreira AK, Matsuo AL, Massaoka MH, et al. (2014) Mastoparan induces apoptosis in B16F10-Nex2 melanoma cells via the intrinsic mitochondrial pathway and displays antitumor activity in vivo. Peptides 68: 113-119. [Crossref]
- Yamada Y, Shinohara Y, Kakudo T, Chaki S, Futaki S, et al. (2005) Mitochondrial delivery of mastoparan with transferrin liposomes equipped with a pH-sensitive fusogenic peptide for selective cancer therapy. Int J Pharm 303: 1–7. [Crossref]
- Jones S, Howl J (2004) Charge delocalisation and the design of novel mastoparan analogues: enhanced cytotoxicity and secretory efficacy of [Lys5, Lys8, Aib10]MP. Regul Pept 121: 121–128. [Crossref]
- Ruoslahti E, Pierschbacher MD (1987) New perspectives in cell adhesion: RGD and integrins. Science 238: 491-497. [Crossref]
- Ribeiro SP, Mendes MA, Santos LD, Souza BM, Marques MR, et al. (2004) Structural and functional characterization of N-terminally blocked peptides isolated from the venom of the social wasp Polybia paulista. Peptides 25: 2069–2078. [Crossref]
- Mendes MA, de Souza BM, Palma MS (2005) Structural and biological characterization of three novel mastoparan peptides from the venom of the neotropical social wasp Protopolybia exigua (Saussure). Toxicon; 45: 101–106. [Crossref]
- Wang K, Zhang B, Zhang W, Yan J, Li J, et al. (2008) Antitumor effects, cell selectivity and structure-activity relationship of a novel antimicrobial peptide polybia-MPI. Peptides 29: 963–968. [Crossref]
- de Souza BM, da Silva AV, Resende VM, Arcuri HA, Dos Santos Cabrera MP, et al. (2009) Characterization of two novel polyfunctional mastoparan peptides from the venom of the social wasp Polybia paulista. Peptides 30: 1387-1395. [Crossref]
- Ward PS (1998) Systematics, biogeography and host plant associations of the Pseudomyrmex viduus group (Hymenoptera: Formicidae), Triplaris- and Tachigali-inhabiting ants. Zool J Linn Soc 126: 451-540.
- Hink WF, Pappas PW, Jaworski DC (1994) Partial biochemical characterization of venom from the ant, Pseudomyrmex triplarinus. Toxicon 32: 763-772. [Crossref]
- Pan J, Hink WF (2000) Isolation and characterization of myrmexins, six isoforms of venom proteins with anti-inflammatory activity from the tropical ant, Pseudomyrmex triplarinus. Toxicon 38: 1403-1413. [Crossref]
- Hink WF, Butz JJ (1985) Primary culture of ant venom gland cells. In Vitro Cell Dev Biol 21: 333-339. [Crossref]
- Schultz DR, Arnold PI (1984) Immunochemical and clinical studies of venom from the ant Pseudomyrmex sp. In: Tu AT (ed). Handbook of Natural Toxicon, Insect Poisons, Allergens and Other Invertebrate Venoms 243-290. Marcel Dekker, New York, USA.
- Altman RD, Schultz DR, Collins-Yudiskas B, Aldrich J, Arnold PI, et al. (1984) The effects of a partially purified fraction of an ant venom in rheumatoid arthritis. Arthritis Rheum 27: 277-284. [Crossref]
- Jetter K, Hamilton J, Klotz J (2002) Eradication costs calculated: red imported fire ants threaten agriculture, wildlife and homes. Calif Agr 56: 26-34.
- Baer H, Liu TY, Anderson MC, Blum M, Schmid WH, et al. (1979) Protein components of fire ant venom (Solenopsis invicta). Toxicon 17: 397-405. [Crossref]
- Howell G, Butler J, Deshazo RD, Farley JM, Liu HL, et al. (2005) Cardiodepressant and neurologic actions of Solenopsis invicta (imported fire ant) venom alkaloids. Ann Allergy Asthma Immunol 94: 380-386. [Crossref]
- Arbiser JL, Kau T, Konar M, Narra K, Ramchandran R, et al. (2007) Solenopsin, the alkaloidal component of the fire ant (Solenopsis invicta), is a naturally occurring inhibitor of phosphatidylinositol-3-kinase signaling and angiogenesis. Blood 109: 560-565. [Crossref]
- Karlsson I, Zhou X, Thomas R, Smith AT, Bonner MY, et al. (2015) Solenopsin A and analogs exhibit ceramide-like biological activity. Vasc Cell 7: 5. [Crossref]
- Kauffmann-Zeh A1, Rodriguez-Viciana P, Ulrich E, Gilbert C, Coffer P, et al. (1997) Suppression of c-Myc-induced apoptosis by Ras signalling through PI(3)K and PKB. Nature 385: 544-548. [Crossref]
- Coffer PJ, Jin J, Woodgett JR (1998) Protein kinase B (c-Akt): a multifunctional mediator of phosphatidylinositol 3-kinase activation. Biochem J 335: 1-13. [Crossref]
- Aoki M, Batista O, Bellacosa A, Tsichlis P, Vogt PK (1998) The akt kinase: molecular determinants of oncogenicity. Proc Natl Acad Sci U S A 95: 14950-14955. [Crossref]
- Gerber HP, McMurtrey A, Kowalski J, Yan M, Keyt BA, et al. (1998) Vascular-endothelial growth factor regulates endothelial cell survival through the phosphatidylinositol 3’-kinase/Akt signal transduction pathway: requirement for Flk-1/KDR activation. J Biol Chem 273: 30336-30343. [Crossref]
- Dhawan P, Singh AB, Ellis DL, Richmond A (2002) Constitutive activation of Akt/protein kinase B in melanoma leads to up-regulation of nuclear factor kappa B and tumor progression. Cancer Res 62: 7335-7342. [Crossref]
- Kulik G, Klippel A, Weber MJ (1997) Antiapoptotic signalling by the insulin-like growth factor I receptor, phosphatidylinositol 3-kinase, and Akt. Mol Cell Biol 17: 1595-1606. [Crossref]
- Chee KG, Longmate J, Quinn DI, Chatta G, Pinski J, et al. (2007) The AKT inhibitor perifosine in biochemically recurrent prostate cancer: a phase II California/Pittsburgh cancer consortium trial. Clin Genitourin Cancer 5: 433-437. [Crossref]
- Kaufman M (2013) Phase III trial of perifosine in multiple myeloma halted. Downloaded from http://www.cancernetwork.com/multiple-myeloma/phase-iii-trial-perifosine-multiple-myeloma-halted. Accessed August 22, 2016.
- Ahn MY, Han JW, Hwang JS, Yun EY, Lee BM (2014) Anti-inflammatory effect of glycosaminoglycan derived from Gryllus bimaculatus (a type of cricket, insect) on adjuvant-treated chronic arthritis rat model. J Toxicol Environ Health A 77: 1332-1345. [Crossref]
- Hahn BS, Cho SY, Ahn MY, Kim YS (2001) Purification and characterization of a plasmin-like protease from Tenodera sinensis (Chinese mantis). Insect Biochem Mol Biol 31: 573-581. [Crossref]
- Latifi M, Alikhani MY, Salehzadeh A, Nazari M, Bandani AR, et al. (2015) The antibacterial effect of American cockroach hemolymph on the nosocomial pathogenic bacteria. Avicenna J Clin Microbiol Infect 2: e23017.
- Gasch T, Schott M, Wehrenfennig C, Düring RA, Vilcinskas A (2013) Multifunctional weaponry: the chemical defenses of earwigs. J Insect Physiol 59: 1186-1193. [Crossref]
- Dive G, Bouillon C, Sliwa A, Valet B, Verlaine O, et al. (2013) Macrocycle-embedded β-lactams as novel inhibitors of the Penicillin Binding Protein PBP2a from MRSA. Eur J Med Chem 64: 365-376. [Crossref]
- Jin X, Mei H, Li X, Ma Y, Zeng AH, et al. (2010) Apoptosis-inducing activity of the antimicrobial peptide cecropin of Musca domestica in human hepatocellular carcinoma cell line BEL-7402 and the possible mechanism. Acta Biochim Biophys Sin (Shanghai) 42: 259–265. [Crossref]
- Sherman RA (2014) Mechanisms of maggot-induced wound healing: what do we know, and where do we go from here? Evid Based Complement Alternat Med 2014: 592419. [Crossref]
- Hammond E (2015) Amid controversy and irony, Costa Rica’s INBio surrenders biodiversity collections and lands to the state. TWN Third World Network. Downlaoded from http://www.twn.my/title2/biotk/2015/btk150401.htm. Accessed August 22, 2016.
- Schumacher MJ, Schmidt JO, Egen NB (1989) Lethality of 'killer' bee stings. Nature 337: 413. [Crossref]
- Cragg GM, Newman DJ (2013) Natural products: a continuing source of novel drug leads. Biochim Biophys Acta 1830: 3670-3695. [Crossref]
- Collen B, Böhm M, Kemp R, Baillie JEM (2012) Spineless: status and trends of the world’s invertebrates. Zoological Society of London, UK.
- Costa-Neto EM (2005) Entomotherapy, or the medicinal use of insects. J Ethnobiol 25: 93-114.
- Feng Y, Zhao M, He Z, Chen Z, L Sun (2009) Research and utilization of medicinal insects in China. Entomol Res 39: 313–316.
- Meyer-Rochow VB (2013) Ethno-entomological observations from North Korea. J Ethnobiol Ethnomed 9:7.