Abstract
Potassium equilibration between the intracellular cytosol and extracellular space, which is regulated by the Na+/K+-ATPase sodium pump, is associated with the generation of resting membrane potential. It is also important for generating and repolarizing action potentials in excitable cells. Equilibrium can easily vary with alterations in extracellular (i.e. plasma) potassium levels. Here, we investigated the influence of potassium level reduction in non-excitable and excitable cells by ouabain treatment, which suppresses Na+/K+-ATPase activity, on cell proliferation ability and viability. An intracellular decrease in potassium level was linked to a mitochondrial potassium level decrease. Although intracellular and mitochondrial potassium levels decreased with ouabain treatment in non-excitable, undifferentiated P19 cells, cell death and growth suppression were not observed after exposure. In contrast, in primary neurons—excitable cells—a decrease in intracellular potassium concentration by ouabain treatment led to cell death. These findings suggest that Na+/K+-ATPase activity is fundamentally not necessary for the proliferation of non-excitable cells but has a direct effect on the viability of excitable cells.
Keywords
intracellular potassium level, mitochondrial depolarization, Na+/K+-ATPase, non-excitable cell, ouabain
Abbreviations used
a-MEM, alpha minimum essential medium; DiSC3(5), 3,3¢-Dipropylthiacarbocyanine iodide; MTT, 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl-tetrazolium bromide; Na+/K+-ATPase, sodium-potassium pump; PTP, permeability transition pore; ROS, reactive oxygen species
Introduction
Normal blood potassium levels are regulated in quite a narrow range, between 3.5 and 5.0 mM (or mEq/L) [1]. Causes of dehydration, such as gastrointestinal disorders (including diarrhea), renal failure, and excessive perspiration, can disturb potassium homeostasis [1]. Both hyperkalemia and hypokalemia can cause muscle dysfunction [1-4]. The former cause’s tachycardia and the latter bradycardia, and both carry a risk of cardiac arrest [1-3]. Because potassium ions are at their highest concentration in muscles [1,4], changes in potassium levels can lead directly to myopathy. Rarely, hyperkalemia or hypokalemia can manifest as epilepsy [5,6], sensorineural deafness, ataxia, and mental retardation [7]. Recent reports have demonstrated that a reduction in intracellular potassium ion levels has protective effects on excitotoxicity or neurodegenerative events in primary neurons. [8-10]. The importance of potassium homeostasis is therefore worth reviewing to increase our understanding of these aspects of neurophysiology.
Most potassium ions in the body are found inside the cells. Intracellular potassium levels are maintained at about 140 mM higher than in the extracellular spaces, where they are present at about 3.5 to 5.0 mM [1]. Concentration gradients of potassium ions are basically formed by a sodium-potassium pump (Na+/K+-ATPase), which pumps potassium ions into the cell and pumps sodium ions out of the cell [1,11]. When the pump is inhibited by a cardiac glycoside such as ouabain, sodium ions in the cardiomyocytes, which are translocated into the cells during depolarization, cannot be pumped out. The increased intracellular sodium ion content inhibits the activity of the sodium–calcium exchanger, perpetuating high intracellular calcium levels and thus sustaining myocardial contraction [11]. Inhibition of Na+/K+-ATPase, for example, by cardiac glycosides, also reduces intracellular potassium ion levels.
Potassium gradients are critically important for many physiological processes, including maintenance of cellular membrane potential and generation and transduction of action potentials in muscles and neurons [11,12]. The level of membrane potential is determined by this gradient and is calculated by using Nernst’s equation [1]. Opening of the voltage-gated potassium channel is responsible for repolarization of the action potential. Promotion of channel opening shortens the refractory period of the action potential, whereas suppression of channel opening prolongs it [1,3]. Unlike in non-excitable cells, this potassium gradient may exert an indirect protective effect on excitable cells. Vascular wall collapse after cerebral ischemic insult severely alters the levels of electrolytes—not only calcium ions but also extracellular potassium ions—in the brain parenchyma, and this can result in nerve cell injury. Astrocytes incorporate both extracellular calcium ions and extracellular potassium ions and exert protective effects on nerve cells [13].
Potassium ion levels are lower in the mitochondrial matrix than in the cytoplasm. In a way similar to that in the plasma membrane, the mitochondrial inner membrane restricts ion transport between the mitochondrial matrix and the cytoplasm; ions can permeate through the membrane only when specific channels are open or when transport proteins namely pumps or carriers are operating [14]. Proton gradients form the membrane potential of the mitochondrial inner membrane. Because the electron transport chain pumps out protons from the mitochondrial matrix, the mitochondrial membrane potential is negative at about –180 mV [14]. Therefore, flow of cations, including potassium ions, into the matrix through open channels causes mitochondrial depolarization [14-17]. Intracellular calcium ion overload induces the opening of mitochondrial permeability transition pore (PTP), allowing a variety of ions and cellular components with molecular weights below 1200 Da to permeate from the mitochondrial matrix to the cytosol. Permeation of protons through PTP causes depolarization of the mitochondrial membrane and uncoupling of the mitochondrial oxidative phosphorylation system, as well as mitochondrial morphological changes such as swelling. These alterations induce the production of reactive oxygen species (ROS) and apoptotic proteins such as cytochrome C and inhibit the synthesis of the cellular fuel ATP [15-17]. Furthermore, ROS and calcium ions enhance the permeability of the mitochondrial membrane, and influx of potassium ions into the matrix also triggers the opening of PTP [18]. It is therefore possible that the opening of mitochondrial membrane potassium channels and mitochondrial dysfunction are correlated.
Fluorescent indicators are available for investigating various features of potassium homeostasis. Asante Potassium Green is useful for detecting intracellular potassium levels [19,20]. The cationic fluorescent dye 3,3¢-dipropylthiacarbocyanine iodide (DiSC3(5)) is an indicator of membrane potential; it accumulates in the mitochondria when the inner mitochondrial membrane is depolarized [21]. Valinomycin is a potassium ionophore and could thus induce mitochondrial depolarization. Recently, we used DiSC3(5) to measure the membrane potential of mitochondria in cultured neurons found that the extent of depolarization and degree of neuronal cell death were correlated [8].
Here, we measured intracellular potassium levels with fluorescent indicators and found that the extent of mitochondrial depolarization seemed to be influenced by the intracellular potassium level. We also found that, when intracellular potassium levels were decreased by ouabain treatment, neurons (i.e. excitable cells) degenerated, whereas non-excitable cells proliferated normally. These findings further elucidate the physiological significance of increases and decreases in potassium ion levels in cells.
Materials and Methods
Chemicals
DiSC3(5) was purchased from Anaspec Inc. (San Jose, CA). Ouabain was obtained from Sigma-Aldrich Corp. (St. Louis, MO). Hanks’s balanced salt solution without Ca2+ or Mg2+, together with 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl-tetrazolium bromide (MTT) and valinomycin, was obtained from Nacalai Tesque (Kyoto, Japan). Asante Potassium Green-2 was obtained from TEFLabs, Inc (Austin, TX). All other chemicals were of standard grade.
P19 cell line
The P19 strain of embryonal carcinoma cells was obtained from the American Type Culture Collection (Manassas, VA). P19 cells were maintained at 37°C in 5% CO2 in a humidified atmosphere in alpha minimum essential medium (a-MEM) supplemented with 10% fetal bovine serum. This cell line can be induced to differentiate into neural cells by treatment with all-trans-retinoic acid [22] or into cardiac muscle cells by exposure to dimethyl sulfoxide [23]. Here, however, we investigated the proliferation of undifferentiated cells by division. Cells were plated at a density of 60,000 cells/well of 24-well plates and passed every 48 hours since they reached approximately confluence within 48 h after every passage.
Primary neurons
The protocol used here met the guidelines of the Japanese Society for Pharmacology and was approved by Setsunan University’s Committee for Ethical Use of Experimental Animals. Pregnant Std-ddY mice were housed in metallic breeding cages with a light–dark cycle of 12 h–12 h and a humidity of 55% at 23°C and given free access to food and water. Primary cultures of cortical neurons were prepared from 15-day-old embryonic ddY mice, as originally described by di Porzio et al. [24], with some modifications. In brief, the dissected neocortex was incubated for 10 min at room temperature in 0.02% (wt./vol.) EDTA solution. After removal of the medium, the cells were suspended by gentle trituration in Neurobasal medium (Thermo Fisher Scientific; Waltham, MA) supplemented with NeuroBrew-21 (Miltenyi Biotec K.K.; Bergisch Gladbach, Germany), 100 U/mL penicillin, and 100 μg/mL streptomycin. After being centrifuged at 500 g and then resuspended in Neurobasal medium, the cortical neurons were seeded onto poly-L-lysine-coated dishes and incubated at 37°C in 95% air – 5% CO2. After 2 days in vitro, the cells were treated with cytosine-β-D-arabinofuranoside at 5 μM for 24 h to minimize the growth of proliferative contaminants such as glial cells. The culture medium was replaced with fresh Neurobasal medium every 3 days. At 8 to 10 days in vitro, the cells were used in the observations described below.
Observation of valinomycin-induced mitochondrial depolarization by using DiSC3(5)
Recording medium was prepared using Hank’s balanced salt solution without Ca2+ or Mg2+; the medium contained 137 mM NaCl, 4.17 mM NaHCO3, 0.34 mM Na2HPO4, 5.37 mM KCl, 0.44 mM KH2PO4, and 5.55 mM D-glucose and was supplemented with 20 mM HEPES-NaOH (pH 7.5), 2.0 mM CaCl2, and 250 nM DiSC3(5). This fluorescent dye has been used as an indicator for monitoring the depolarization of isolated mitochondria [21]. The culture media were replaced with the recording medium and cells were incubated for 30 min at 37°C in 95% air – 5% CO2. Fluorescence emission was monitored at an excitation wavelength of 635 nm and an emission spectrum from 650 to 700 nm. Photos or photo bursts were taken under a FluoView FV1000 confocal microscope (Olympus, Tokyo, Japan). In the photo burst experiments, 38 photos were taken every 5 s for a total of 180 s. The first 30 s was shot as background, and then the valinomycin at the final concentration of 2 aM was added. The intensity of fluorescence emission in the region of interest was determined by using the free application Image J (National Institutes of Health; Bethesda, MD) for quantitative analysis.
Observation of intracellular potassium levels by using Asante Potassium Green-2
Recording medium was prepared by using Hanks’s balanced salt solution without Ca2+ or Mg2+; the medium contained 137 mM NaCl, 4.17 mM NaHCO3, 0.34 mM Na2HPO4, 5.37 mM KCl, 0.44 mM KH2PO4, and 5.55 mM D-glucose and was supplemented with 20 mM HEPES-NaOH (pH 7.5), 2.0 mM CaCl2, and 100 nM Asante Potassium Green-2, which is a fluorescent dye for potassium ions. P19 cells were cultured with a concentration of ouabain for 24 h and were then stained with Asante Potassium Green-2 by incubating with the recording medium for 30 min at 37°C in 95% air – 5% CO2. Fluorescence emission was then monitored at an excitation wavelength of 488 nm and an emission spectrum from 520 to 560 nm. Photos were taken under a FluoView FV1000 confocal microscope. The intensity of fluorescence emission in the region of interests in this case the whole area of each photo was determined by using the free application Image J for quantitative analysis.
MTT assay
P19 cells were cultured in a-MEM (as described above) with or without ouabain for up to 72 h at 37°C in 95% air – 5% CO2. In the case of Figure 1, the medium of P19 cells cultured in a-MEM (as described above) for 24 h was replaced by and incubated in Hanks’s balanced salt solution without Ca2+ or Mg2+; the medium contained 137 mM NaCl, 4.17 mM NaHCO3, 0.34 mM Na2HPO4, 5.37 mM KCl, 0.44 mM KH2PO4, and 5.55 mM D-glucose and was supplemented with 20 mM HEPES-NaOH (pH 7.5) and 2.0 mM CaCl2 with or without ouabain for 30 min at 37°C in 95% air – 5% CO2. Primary neurons were incubated with ouabain for 30 min at 37°C under an atmosphere of 95% air – 5% CO2; the incubation medium was then replaced with fresh Neurobasal medium (as described above) without ouabain. This was followed by additional incubation for 24 h at 37°C under an atmosphere of 95% air – 5% CO2. After the incubations of both types of cells under different conditions, the medium was exchanged for a solution containing 0.5 mg/mL MTT and 33 mM glucose in phosphate-buffered saline; this was followed by further incubation for 2 h at 37°C under an atmosphere of 95% air – 5% CO2. The resulting blue formazan dye an indicator of living cells—was solubilized with 0.04 mol/L HCl in 2-propanol to measure the optical density by a microplate reader Model 680 (Biorad; Hercules, CA) at a wavelength 570 nm.
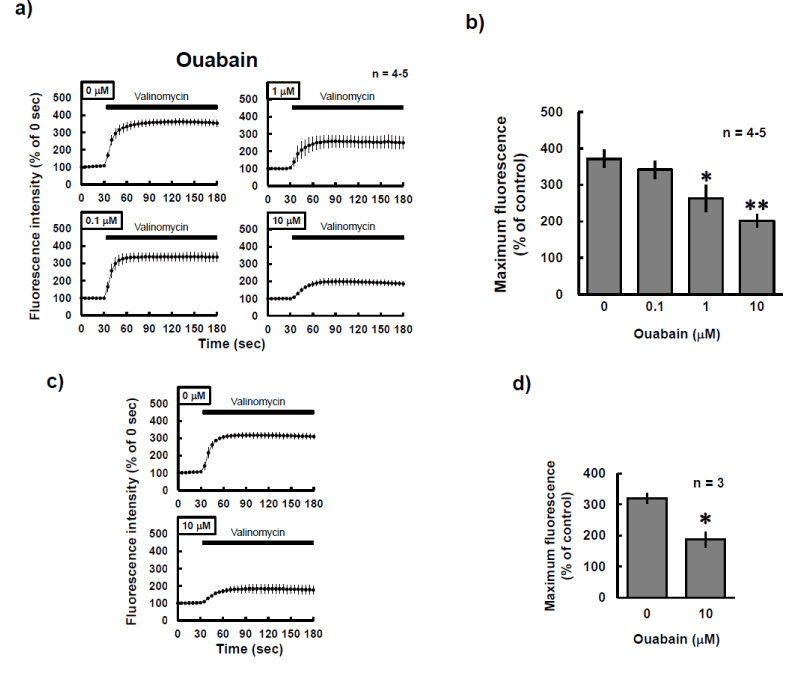
Figure 1: Effects of ouabain on valinomycin-induced alteration of DiSC3(5) fluorescence in P19 cells. P19 cells were cultured at the indicated concentrations of ouabain for a, b) 24 h or c, d) 30 min. a, c) Cells were then loaded with 250 nM DiSC3(5) for 30 min at 37ºC and observed by confocal microscopy, with fluorescence emission at a wavelength of 675 nm. During photo-burst experiments, 2 aM valinomycin was added. The fluorescence intensity of the region of interest was quantified by using Image J. Horizontal black bar indicates period of exposure to valinomycin. b, d) Maximum fluorescence values during continuous shooting experiments in a) or c) were calculated and compared. The experiments were performed several times, as indicated. b) *P<0.05, **P<0.01 vs. control experiment (Dunnett’s test). d) *P<0.05 vs. control experiment (Welch’s t-test).
Data analysis
Fluorescence intensities of photos were quantified by using Image J. Each region of interest surrounded a region of whole neurons, except in the case of nonspecific regions, which always had high signal intensities. All data were expressed as means ± S.E., and statistical significance was determined by using Dunnett’s test or Welch’s t-test.
Results
Valinomycin-induced mitochondrial depolarization can be estimated by using the indicator DiSC3(5)
DiSC3(5), an indicator of mitochondrial membrane potential, was incubated with P19 cells for 30 min to determine fluorescence changes after addition of the potassium ionophore valinomycin. Figure 2a is typical fluorescence image of P19 cells before (upper panel) and after (lower panel) exposure to the valinomycin at 2 aM for 150 s. The ionophore induced a markedly sustained increase in fluorescence intensity (Figure 2b). During culture, the MTT reduction ability of P19 cells increased substantially with time (Figure 3a) and the cells seemed to reach confluence within 48 h after the start of each passage. Also, at each point in the culture, valinomycin induced an increase in DiSC3(5) fluorescence to about the same level (Figures 3b and c).
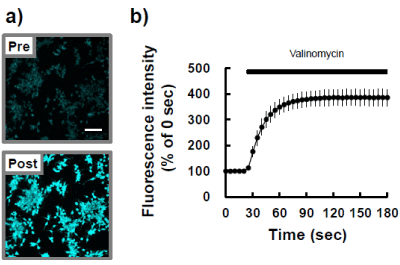
Figure 2: Typical DiSC3(5) fluorescence in P19 cells. P19 cells were loaded with 250 nM DiSC3(5) for 30 min at 37ºC and then observed by confocal microscopy, with fluorescence emission at a wavelength of 675 nm. During photo-burst experiments, 2 aM valinomycin was added. a) Representative photomicrographs of pre- and post-treatment with valinomycin are shown. b) The fluorescence intensity of the region of interest was quantified by using Image J. Horizontal black bar indicates period of exposure to valinomycin.
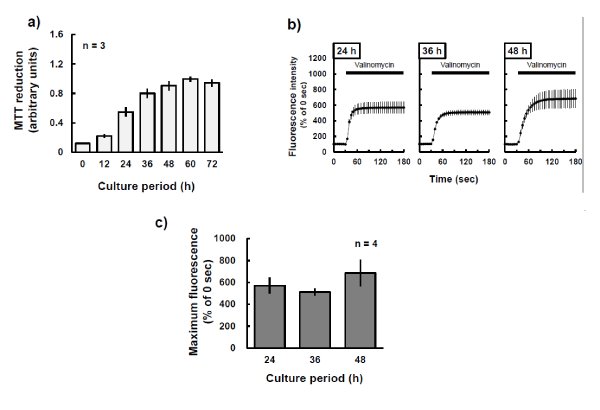
Figure 3: Consistent response to valinomycin during proliferation of P19 cells. a) P19 cells were cultured for the periods indicated and the ability of their mitochondria to reduce MTT were estimated. The experiments were performed three times. b) Cells at the indicated culture times were loaded with 250 nM DiSC3(5) for 30 min at 37ºC and then observed by confocal microscopy, with fluorescence emission at a wavelength of 675 nm. During photo-burst experiments, 2 aM valinomycin was added. The fluorescence intensity of the region of interest was quantified by using Image J. Horizontal black bar indicates period of exposure to valinomycin. c) The maximum fluorescence values during photo-burst experiments in b) were calculated and compared. The experiments were performed four times. No significant differences were found among culture periods (Dunnett’s test).
Reduction of intracellular potassium levels suppresses valinomycin-induced mitochondrial depolarization
Intracellular potassium concentrations can be detected with the indicator Asante Potassium Green-2. The fluorescence intensity of Asante Potassium Green-2 was weakened in a concentration-dependent manner by treatment of P19 cells for 24 h with ouabain, an inhibitor of Na+/K+-ATPase (Figure 4). We hypothesized that the low intracellular potassium levels induced by ouabain would result in a weak response to valinomycin-induced depolarization of the mitochondria. Indeed, treatment with ouabain for 24 h revealed a concentration-dependent decrease in maximum fluorescence after treatment with valinomycin in P19 cells loaded with DiSC3(5) (Figures 1a and b). These findings suggested that valinomycin-induced mitochondrial depolarization was weakened by the reduction of intracellular potassium levels. The decrease in valinomycin-induced depolarization was also observed even after a 30-min exposure to ouabain (Figure 1c). This suggested that the decline in intracellular potassium levels was completed within 30 min after the start of exposure to ouabain and that the effect continued for at least 24 h.
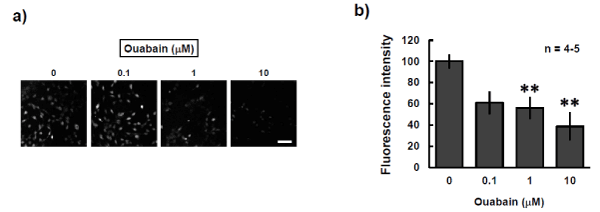
Figure 4: Effect of ouabain on intracellular potassium levels in P19 cells. P19 cells were cultured with the indicated concentrations of ouabain for 24 h and were then stained with Asante Potassium Green-2. a) Representative photo images of P19 cells stained with Asante after the 24-h ouabain exposure period are shown. Scale bar = 100 am. b) The fluorescence intensities of the regions of interest were quantified and compared. The experiments were performed four or five times. **P<0.01 vs. control experiment (Dunnett’s test).
Although ouabain induces neuronal cell death, it does not affect P19 cell-line survival
Whether a decrease in the intracellular potassium concentration affects cell survival appeared to be determined by whether the cells were excitable or non-excitable. Interestingly, after 24 h of ouabain exposure, the population density of P19 cells did not differ from that of control cultures (Figure 5). Even after 36 h of culture with the ouabain concentration increased to 100 aM, the cells continued to grow similarly to the controls. In primary neurons, as in P19 cells, ouabain treatment for 30 min dose-dependently weakened valinomycin-induced mitochondrial depolarization (Figure 6a and b), and this was likely due to a reduction in intracellular potassium levels. Even when the neurons were exposed to fresh medium after the 30-min ouabain exposure, they degenerated the day after the exposure in an ouabain-concentration–dependent manner (Figure 6c).
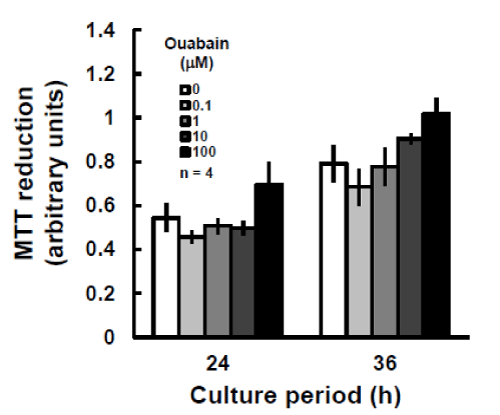
Figure 5: Effects of ouabain on proliferation of P19 cells. P19 cells were cultured with the indicated concentrations of ouabain for 24 or 36 h and the abilities of their mitochondria to reduce MTT were estimated. The experiments were performed four times. There were no significant differences between any of the ouabain exposure groups vs. control groups at each culture time (Dunnett’s test).
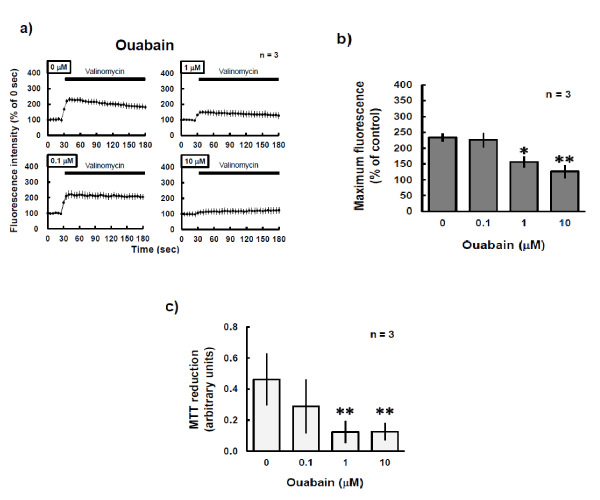
Figure 6: Effects of ouabain on primary cultured neurons. a) Primary neurons prepared from mouse embryos were exposed to ouabain for 30 min at 37ºC during loading with 250 nM DiSC3(5) and then examined by confocal microscopic observation, with fluorescence emission at a wavelength of 675 nm. During photo-burst experiments, 2 aM valinomycin was added. The fluorescence intensity of the region of interest was quantified by using Image J. Horizontal black bar indicates period of exposure to valinomycin. b) Maximum fluorescence values during photo-burst experiments in a) were calculated and compared. The experiments were performed three times. *P<0.05, **P<0.01 vs. control experiment (Dunnett’s test). c) Primary neurons were exposed to ouabain for 30 min. The incubation medium was then replaced with fresh ouabain-free medium, followed by additional incubation for 24 h and subsequent MTT assay. The experiments were performed three times. **P<0.01 vs. control experiment (Dunnett’s test).
Discussion
P19 cells are an embryonic stem cell carcinoma line that can be induced to differentiate into excitable cells such as neurons and cardiomyocytes [22,23]. In the past we have used them to study neuronal stem cells [25], although the cells used here were undifferentiated proliferating P19 cells and were non-excitable. The fact that astrocytes reduce extracellular potassium concentrations through the action of Na+/K+-ATPase in the central nervous system [13,26,27] indicates that it is important to elucidate the mechanism of potassium homeostasis regulation in non-excitable cells. Although endogenous cardiac glycosides have not yet been identified, it has been suggested that they interfere with the function of Na+/K+-ATPase and participate in the onset of hypertension [28]. This evidence supports the need to examine the action of cardiac glycosides on stem cells.
Because valinomycin increases membrane potassium permeability and induces mitochondrial depolarization, the fluorescence intensity of DiSC3(5) loaded into undifferentiated P19 cells was likely increased by valinomycin treatment. The level of valinomycin-induced mitochondrial depolarization did not change during P19 cell proliferation (Figure 3). Therefore, the cells were likely proliferating without differentiation or any notable special change in intracellular potassium levels. Ouabain treatment for 24 h decreased the fluorescence intensity of Asante Potassium Green-2 (Figure 4) and suppressed the maximum valinomycin-induced mitochondrial depolarization (Figures 1a and b) in P19 cells. These findings suggest that the level of valinomycin-induced mitochondrial depolarization reflected the intracellular potassium level. Influx of potassium ions into the matrix leads to calcium influx and opening of PTP [18], but the indicator DiSC3(5) did not show a maximum increase in fluorescence under cell treatment with ouabain at 1 aM or more. As the potassium level in the cytosol declined, the amount of potassium flowing into the matrix decreased, so it was probably not able to reach a state in which PTP could open. This suggests that a decrease in intracellular potassium level might prevent mitochondrial toxicity.
A 30-min incubation period was sufficient for ouabain to exert its effects on P19 cells (Figures 1c and d), whereas this treatment had no effect on the proliferation of P19 cells, even after 24 h (Figure 5). A tendency to promote proliferation was even observed at 100 aM of ouabain. This suggests that the sodium pump is not necessarily required for cell proliferation. The continuing presence of valinomycin-induced mitochondrial depolarization indicated that ouabain treatment did not eliminate the potassium gradient between the cytosol and the matrix. Even in the presence of ouabain, some transporters, including the potassium-proton exchanger [18], could have formed a gradient. Similarly, the potassium leak channel and transporters such as potassium-chloride co-transporters or sodium-potassium-chloride co-transporters [1,27] on the plasma membrane might provide an artificial potassium gradient between the extra- and intracellular spaces in P19 cells.
In contrast to the results in P19 cells, ouabain treatment of primary neurons for 30 min was lethal in a concentration-dependent manner (Figure 6), and this likely occurred in response to a decrease in intracellular potassium level (data not shown). A transient decrease in intracellular potassium levels induces hyperpolarization in quiescent cells, whereas inhibition of the sodium pump for a long time produces new artificial potassium equilibrium. At this time, the absolute value of the membrane potential is small (i.e. the membrane is in depolarization) and an action potential is readily generated [1,27]. Spontaneous action potentials may be generated under culture conditions. Delayed repolarization of those action potentials can cause extreme excitation of culture neurons, and this is probably harmful. In our previous work [8], conditions that reduced the fluorescence intensity of DiSC3(5) were brought about by opening of the G-protein-coupled inwardly rectifying potassium channels, and this had a protective effect on the culture neurons. We consider that this observation resulted from a decrease in intracellular potassium levels, although it could not be reproduced by the inhibition of Na+/K+-ATPase in our current study. In the central nervous system, extracellular potassium ions likely increase the excitability of nerve cells, and astrocytes protect the neurons by incorporating potassium ions themselves [13,27]. Astrocytes probably take up extracellular potassium ions by increasing the activity of the Na+/K+-ATPase pump by a mechanism that depends on an increase in intracellular calcium ion levels or the activation of adrenergic β receptors, or both [27]. Potassium channel openers expressed on the cell membrane and mitochondrial inner membrane, such as minoxidil and nicorandil, have a neuronal protective effect [9,10]. In the future, we will need to examine the fluctuation of potassium homeostasis and the neuroprotection associated with potassium channel opening on the cell membrane.
The fluorescent indicator Asante Potassium Green-2 used here was suitable for measuring potassium levels in cells in the resting state, but not for measuring decreases in potassium levels by tracking over time, because it did not fade in seconds, probably because of a high affinity for, and weak dissociation from, potassium. However, it is possible to measure increases in intracellular potassium levels kinetically by adding potassium channel openers and instantaneously increasing extracellular potassium levels, although increasing intracellular potassium levels is not realistic because it rarely occurs in normal cells. Therefore, here, we measured valinomycin-induced mitochondrial depolarization over a period of seconds as a substitute for direct measurement of intracellular potassium ion levels. Because a method of measuring the fluorescence of a thallium ion indicator has now been invented by utilizing the fact that thallium ions penetrate potassium ion channels [29], we are now investigating whether it is useful to investigate the potassium homeostasis of primary culture neurons.
Plasma potassium concentrations depend in part on food intake [1-4]. Unlike over a short period, over a prolonged period excessive (or reduced) intake of potassium can cause excitable cells to be (or not to be) sensitive to excitation. Factors that modulate the equilibrium of potassium ions from outside the cell to inside are insulin, β adrenergic transmission, and the state of acid–base equilibrium [1,30]. Therefore, intracellular potassium concentrations tend to decrease in insulin-deficient type I diabetes and, conversely, insulin replacement therapy can raise intracellular potassium concentrations. Adrenergic β receptor agonists induce potassium transfer into the cell, whereas β blockers lower intracellular potassium levels [27,31]. Adrenergic β-receptor agonists cause astrocytes take up extracellular potassium ions by increasing sodium pump activity [4,27]. Huge numbers of patients are taking these types of drugs for diabetes (insulin replacement therapy), asthma (β2 agonists), and hypertension (β1 antagonists). It is undeniable that neurons in the central nervous system are potentially vulnerable to these drugs.
Our findings indicate that, irrespective of whether cells are non-excitable or excitable, it is important to elucidate the importance of potassium homeostasis and its disturbance in the mechanism of dysfunction of excitable cells. This information will lead to the development of completely new therapeutic agents for central nervous system diseases.
References
- Weiss JN, Qu Z, Shivkumar K (2017) Electrophysiology of Hypokalemia and Hyperkalemia. Circ Arrhythm Electrophysiol 10: pii: e004667. [Crossref]
- El Fassi D and Nielsen G (2013). Hyperkalemia: a clue to the diagnosis of adrenal insufficiency. Circulation 128: 2620-2621. [Crossref]
- Faggioni M, Knollmann BC (2015) Arrhythmia Protection in Hypokalemia: A Novel Role of Ca2+-Activated K+ Currents in the Ventricle. Circulation 132: 1371-1373. [Crossref]
- Clausen T (2010) Hormonal and pharmacological modification of plasma potassium homeostasis. Fundam Clin Pharmacol 24: 595-605. [Crossref]
- Sakemi T, Ikeda Y, Rikitake O (1996) Tonic convulsion associated with sinus arrest due to hyperkalemia in a chronic hemodialysis patient. Nephron 73: 370-371. [Crossref]
- Vanpee D, Dive A, Ossemann M, Gillet JB (2001) Refractory seizure with hypokalaemia. Postgrad Med J 77: 209-213. [Crossref]
- Scholl UI, Choi M, Liu T, Ramaekers VT, Häusler MG, et al. (2009) Seizures, sensorineural deafness, ataxia, mental retardation, and electrolyte imbalance (SeSAME syndrome) caused by mutations in KCNJ10. Proc Natl Acad Sci U S A 106: 5842-5847. [Crossref]
- Kinjo T, Ashida Y, Higashi H, Sugimura S, Washida M, et al. (2017) Alleviation by GABAB Receptors of Neurotoxicity Mediated by Mitochondrial Permeability Transition Pore in Cultured Murine Cortical Neurons Exposed to N-Methyl-D-aspartate. Neurochem Res 1-10. [Crossref]
- Chen YF, Chen LH, Yeh YM, Wu PY, Chen YF, et al. (2017) Minoxidil is a potential neuroprotective drug for paclitaxel-induced peripheral neuropathy. Sci Rep 7: 45366. [Crossref]
- Kong J, Ren G, Jia N, Wang Y, Zhang H, et al. (2013) Effects of nicorandil in neuroprotective activation of PI3K/AKT pathways in a cellular model of Alzheimer’s disease. Eur Neurol 70: 233-241. [Crossref]
- Iwamoto T, Kita S (2006) Hypertension, Na+/Ca2+ exchanger, and Na+, K+-ATPase. Kidney Int 69: 2148-2154. [Crossref]
- Kovesdy CP (2015) Management of Hyperkalemia: An Update for the Internist. The American Journal of Medicine 128: 1281-1287. [Crossref]
- Leis JA, Bekar LK, Walz W (2005) Potassium homeostasis in the ischemic brain. Glia 50: 407-416. [Crossref]
- Szewczyk A, Wojtczak L (2002) Mitochondria as a pharmacological target. Pharmacol Rev 54: 101-127. [Crossref]
- Halestrap AP, Doran E, Gillespie JP, O’Toole A (2000) Mitochondria and cell death. Biochem Soc Trans 28: 170-177. [Crossref]
- Bernardi P (1992) Modulation of the mitochondrial cyclosporin A-sensitive permeability transition pore by the proton electrochemical gradient. Evidence that the pore can be opened by membrane depolarization. J Biol Chem 267: 8834-8839. [Crossref]
- Bernardi P, Broekemeier KM, Pfeiffer DR (1994) Recent progress on regulation of the mitochondrial permeability transition pore; a cyclosporin-sensitive pore in the inner mitochondrial membrane. J Bioenerg Biomembr 26: 509-517. [Crossref]
- Hawrysh PJ, Buck LT (2013) Anoxia-mediated calcium release through the mitochondrial permeability transition pore silences NMDA receptor currents in turtle neurons. J Exp Biol 216: 4375-4387. [Crossref]
- Rimmele TS, Chatton JY (2014) A novel optical intracellular imaging approach for potassium dynamics in astrocytes. PLoS One 9: e109243. [Crossref]
- Bazzigaluppi P, Dufour S, Carlen PL (2015) Wide field fluorescent imaging of extracellular spatiotemporal potassium dynamics in vivo. Neuroimage 104: 110-116. [Crossref]
- Plásek J, Sigler K (1996) Slow fluorescent indicators of membrane potential: a survey of different approaches to probe response analysis. J Photochem Photobiol B 33: 101-124. [Crossref]
- Jones-Villeneuve EM, McBurney MW, Rogers KA, Kalnins VI (1982) Retinoic acid induces embryonal carcinoma cells to differentiate into neurons and glial cells. J Cell Biol 94: 253-262. [Crossref]
- McBurney MW, Jones-Villeneuve EM, Edwards MK, Anderson PJ (1982) Control of muscle and neuronal differentiation in a cultured embryonal carcinoma cell line. Nature 299: 165-167. [Crossref]
- di Porzio U, Daguet MC, Glowinski J, Prochiantz A (1980) Effect of striatal cells on in vitro maturation of mesencephalic dopaminergic neurones grown in serum-free conditions. Nature 288: 370-373. [Crossref]
- Takarada T, Kou M, Hida M, Fukumori R, Nakamura S, et al. (2016) Protective upregulation of activating transcription factor-3 against glutamate neurotoxicity in neuronal cells under ischemia. J Neurosci Res 94: 378-388. [Crossref]
- Ransom CB, Ransom BR, Sontheimer H (2000) Activity-dependent extracellular K+ accumulation in rat optic nerve: The role of glial and axonal Na+ pumps. J Physiol 522: 427-442. [Crossref]
- Hertz L, Song D, Xu J, Peng L, Gibbs ME (2015) Role of the Astrocytic Na(+), K(+)-ATPase in K(+) Homeostasis in Brain: K(+) Uptake, Signaling Pathways and Substrate Utilization. Neurochem Res 40: 2505-2516. [Crossref]
- Lewis LK, Yandle TG, Hilton PJ, Jensen BP, Begg EJ, et al. (2014) Endogenous ouabain is not ouabain. Hypertension 64: 680-683. [Crossref]
- Titus SA, Beacham D, Shahane SA, Southall N, Xia M, et al. (2009) A new homogeneous high-throughput screening assay for profiling compound activity on the human ether-a-go-go-related gene channel. Anal Biochem 394: 30-38. [Crossref]
- Palmer BF, Clegg DJ (2016) Physiology and pathophysiology of potassium homeostasis. Adv Physiol Educ 40: 480-490. [Crossref]
- Mandić D, Nezić L, Skrbić R (2014) Severe hyperkalemia induced by propranolol. Med Pregl 67: 181-184. [Crossref]