Leptin is an adipose-derived hormone that regulates energy balance. Leptin receptors are expressed in extrahypothalamic sites and several reports showed that leptin can influence feeding and locomotor behavior via direct actions on dopaminergic neurons. The leptin deficient mouse (ob/ob) has been used as an animal model of blunted leptin action, and presents with obesity and mild type 2 diabetes. We used ob/ob mice to study the effect of repeated 7-day methamphetamine (METH) administration analyzing locomotion, behavioral sensitization, and somatosensory thalamic mRNA expression of voltage-gated calcium channels and glutamatergic receptors using RT-PCR. We observed reduced METH-mediated responses in ob/ob mice associated with enhanced in mRNA expression of key voltage-gated and glutamate receptors in the somatosensory thalamus. Results described here are important for understanding the control of locomotion and thalamocortical excitability by leptin
methamphetamine, thalamic nuclei, ob/ob mouse
Abbreviations:
METH: Methamphetamine, DA: Dopamine, OB: Obese ob/ob Mouse, TRN: Thalamic Reticular Nucleus, VB: Ventrobasal
Leptin is an adipose-derived hormone [1] known to control appetite and energy expenditure [2]. Leptin-deficient mice presenting a mutation in the ‘obese’ gene (i.e., homozygous ob/ob mice) were described to develop severe obesity after the fifth postnatal week and to exhibit decreased energy expenditure and hyperphagia [2,3]. Leptin receptors are predominantly expressed in the hypothalamus, although they have also been described to be expressed in extrahypothalamic areas like somatosensory thalamic nuclei and the mesolimibic dopamine system [4-6]. It has been found that in ob/ob mice dopamine is required for hyperphagia [7,8] while manifesting hypoactivation of the dopaminergic system [9] that includes disruption in striatal dopamine D2 receptors (D2R) [10].
Methamphetamine (METH) abuse disrupts dopamine neurotransmission and induces neurophysiological and cognitive alterations in humans [11,12]. Our group has described multiple METH-mediated alterations in mouse cortical and sub-cortical areas [13-16], including changes in mRNA levels of membrane receptors and voltage-gated ion channels in medial prefrontal neurons [17]. Repeated exposure to cocaine, a psychostimulant that can also increase DA neurotransmission, enhanced low threshold T-type calcium channel protein levels in mouse somatosensory thalamus neurons [18]. Selective targeting of T-type calcium channels using specific blockers reduced cocaine-mediated hyperlocomotion concomitantly with a reduction in GABAergic neurotransmission onto thalamic ventrobasal nucleus [19], thus suggesting the involvement of somatosensory ventrobasal thalamic nucleus in locomotor alterations.
We hypothesized that leptin acts as a neuromodulator of psychostimulant-mediated changes in locomotor activity and thalamocortical excitability mediated by the alteration changes in mRNA expression of key voltage-gated calcium channels (P/Q-type and T-type), hyperpolarization-activated cyclic nucleotide-gated channels (HCN2), and glutamate receptor subunits (AMPA and NMDA) [16-19]. Behavioural analyses described that both genotypes (wildtype and ob/ob) showed METH-induced locomotor sensitization (increased locomotion after 7-days compared to day 1) but with lower raw values for ob/ob. In addition, METH-treated ob/ob mouse thalamic tissue showed altered mRNA expression of voltage-gated calcium channels, hyperpolarization-activated cyclic nucleotide-gated channels, and glutamate receptor subunits compared to wildtype thalamic neurons. Our results suggest that leptin deficiency can alter transcriptional regulation in the thalamus and influence basal and METH-elicited locomotor behavior.
Animals
We used female C57BL/6JFcen wild type (WT) mice (3-6 months old, Central Animal Facility at University of Buenos Aires, animal protocol #50-2015) or leptin-deficient, homozygous B6.Cg-Lepob/J, obese ob/ob mice (kindly provided by Dr. J.J. Poderoso, (INIGEM, CONICET-UBA) ). Females were housed four per cage in the absence of males, a condition known to maintain them acyclic. Genotyping of ob/ob littermates was determined during the second postnatal week according to Finocchietto et al. [20]. Principles of animal care were in accordance with the ARRIVE guidelines and CONICET (2003), and approved by its authorities using OLAW/ARENA directives (NIH, Bethesda, MD, USA).
Drug treatments
(+)-Methamphetamine hydrochloride (Sigma, St Louis, MO) was administered subcutaneously (sc) once a day for 7 days (1 mg/Kg, calculated as free base, dissolved in sterile saline solution). The METH regimen used in this study was performed according to studies by González et al. [15]. Vehicle groups received the same volume of sterile saline.
Behavioral studies
Mouse locomotor activity was recorded with an automated system (Ethovision XT 7.0, Noldus, The Netherlands) as previously described [15,18,19]. Total distance traveled (cm) was quantified for a total of 5 minutes prior to injections (basal), and 40 minutes following the last injection of METH/vehicle for days 1 and 7 (test). Drug-induced locomotion was expressed as the ratio between locomotion following drug injection and basal locomotion. Locomotor sensitization was calculated as the ratio of relative locomotion after the 7-day treatment compared to day 1.
Real time PCR
Mice were killed 4 days after treatments and their brains were rapidly removed. The somatosensory thalamic complex (ventrobasal+reticular nuclei) was dissected, placed on dry ice, and stored at -70°C in RNAlater for further assays. Total RNA was isolated using TRIZOL reagent (Invitrogen) according to the manufacturer’s protocol. Five hundred nanograms of RNA were treated with DNAseI (Deoxiribonuclease I) and reverse-transcribed in a 20 µL reaction using M-MLV reverse transcriptase and random hexameres. For quantitative real-time PCR (qPCR) primer sets were designed for the specific amplification of murine Cacna1a, Cacna1g, Cacna1i, Hcn2, Gria1, Grin1, and Gapdh as a housekeeping control gene [17]. Each sample was assayed in duplicate using 4 pmol of each primer, 1X SYBR Green Master Mix, and 2-20 ng of cDNA in a total volume of 13 µL. Amplification was carried out in an ABI PRISM 7500 Sequence Detection System.
Statistical analysis
InfoStat software (www.infostat.com.ar) was used for statistical comparisons. Statistics were performed using two-way (genotype and treatment) ANOVA followed by Bonferroni post hoc tests. Differences were considered significant if p<0.05.
Pharmacological reagents
Drugs were purchased from either Sigma (St. Louis, MO; USA) or Tocris (Ellisville, MO; USA). Reagents for RT-PCR were purchased from Qiagen (GermanTown, MD, USA), Applied Biosystems (Foster City, CA, USA) or Thermofisher-Invitrogen (Waltham, MA, USA).
We measured body weight of WT and ob/ob female mice at the beginning (day 1) and end (day 7) of treatment. Table 1 shows that at both day 1 and day 7 ob/ob mice (OB) were significantly heavier than WT littermates. Paired t-test within each group comparing day 1 vs. day 7 showed no treatment effect on body weight.
Table 1. Body weight at the beginning (day 1) and end (day 7) of treatment.
| |
Body weight (gr) |
WT vehicle |
WT METH |
OB vehicle |
OB METH |
Day 1 |
26.7 ± 1.0 |
25.7 ± 1.1 |
60.1 ± 3.0 |
64.8 ± 1.6 |
Day 7 |
26.4 ± 1.0 |
25.0 ± 1.0 |
59.2 ± 2.1 |
63.8 ± 1.4 |
Two-way ANOVA-Bonferroni (N=5-6); day 1: F(3,21)=141.94 p<0.0001; genotype effect F(1,21)=412.87 p<0.0001 comparing WT vs. OB; day 7: F(3,21)=156.33 p<0.0001; genotype effect F(1,21)=454.02 p<0.0001 comparing WT vs. OB).
We started assessing basal locomotor activation of either WT and ob/ob transgenic mice before METH administration at day 1. Obese ob/ob female mice showed substantially diminished locomotion througout the arena compared to wild type mice (Figure 1B, left). After the first METH administration on day 1, both WT and ob/ob mice increased their locomotion (Figure 1B, right), while maintaining differences between genotypes. At day 7, basal locomotor activity showed interactive effects between genotype and treatment, and we found differences betwee WT-vehicle and OB, but no changes in locomotion after after METH between genotypes (Figure 1C, left). METH-mediated stimulation of locomotor activity was significantly higher at day 7 (Figure 1C, right). METH-mediated sensitization (i.e., increased METH-induced motor activation at day 7 administration compared to day 1) was observed in both WT and ob/ob mice (Figure 1D).
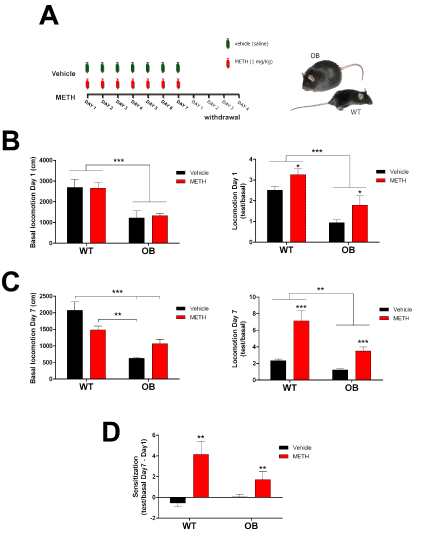
Figure 1. A) Schematic figure showing the METH administration protocol used. Methamphetamine (METH) or vehicle were administered once a day for 7 consecutive days. The ventrobasal thalami were collected 4 days after the last injection. B) Locomotor response at Day 1. Basal (left panel; two-way ANOVA, F(3,21)=8,34 p<0.001; genotype effect F(1,21)=24.97 p<0.0001 comparing WT vs. OB) and drug-induced (right panel; two-way ANOVA, F(3,21)=7.93 p<0.01; genotype effect F(1,21)=19.08 p<0.001, WT vs. OB; treatment effect F(1,21)=5.27 p<0.05, vehicle vs. METH administration) relative locomotion (test/basal) on day 1. C) Locomotor response at Day 7. Basal (left panel; two-way ANOVA, (N=5-6), F(3,21)=16.16 p<0.0001; interaction effect F(1,21)=11.95, p<0.01, Bonferroni post-test among all groups) and drug-induced (right panel; two-way ANOVA, (N=5-6), F(3,21)=13.09 p<0.0001; genotype effect F(1,21)=10,86 p<0.01, WT vs. OB; treatment effect F(1,21)=24.14, p<0.001, vehicle vs. METH administration) relative locomotion (test/basal) on day 7. D) Locomotor sensitization. Relative locomotion day 7 – day1. Two-way ANOVA, (N=5-6), F(3,21)=13.09 p<0.0001; treatment effect F(1,21)=14.21, p<0.01, vehicle vs. METH administration).
We evaluated mRNA expression of voltage-gated calcium channels, hyperpolarization-activated cyclic nucleotide-gated (HCN2) channels, and glutamate receptor subunits in the somatosensory thalamic complex (ventrobasal+reticular nuclei) four days after METH treatment (Figure 2). Assays of P/Q-type Cav2.1 (Cacna1a), and T-type Cav3.3 (Cacna1i) voltage-gated calcium channel subunits showed higher mRNA levels in ob/ob mice relative to WT (Figure 2A,C). No changes were detected for assays of T-type Cav3.1 mRNA levels (Cacna1g) (Figure 2B). The assays of HCN2 showed elevated mRNA levels in ob/ob that were down-regulated by METH (Figure 2D). Assays of NMDA-GluN1 Grin1 (Figure 2E) and AMPA-GluA1 Gria1 (Figure 2F) mRNA levels were observed to be higher in ob/ob mice, independently of METH administration.
Figure 2. mRNA level expression in ventrobasal thalamic nucleus evaluated by RT-PCR. A-C, mRNA level expression of voltage-gated calcium channels subunits Cacna1a (A; P/Q Cav2.1), Cacna1g (B; T Cav3.1), Cacna1i (C; T Cav3.3) (Two-way ANOVA, (N=5), Cacna1a F(3,19)=3.98, p<0.05; genotype effect F(1,19)=6.86, p<0.05 WT vs OB; Cacna1i F(3,19)=3.09, p<0.05; genotype effect F(1,19)=8.33, p<0.05 WT vs OB). D, mRNA level expression of HCN subunits Hcn2 (Two-way ANOVA, F(3,19)=5.00, p<0.05; interaction effect F(1,19)=8.22, p<0.05). E, mRNA level expression of glutamate receptors subunits Gria1 and Grin1 (AMPA GluA1 and NMDA GluN1, respectively; two-way ANOVA, Gria1 F(3,19)=6.06, p<0.01; genotype effect F(1,19)=16.44, p<0.01 WT vs OB; Grin1 F(3,19)=5.29, p<0.05; genotype effect F(1,19)=15.13, p<0.01 WT vs OB;).
Results presented here support the hypothesis that the absence of leptin regulation during development alters psychostimulant-mediated locomotor activity and thalamocortical excitability. In the present study we show that repetitive administration of METH induced lower locomotor stimulation in leptin-null, ob/ob mice. This effect was also observed in the absence of METH. It should be noted that locomotor activity tends to be depressed in obese mice, probably due to abnormal neuroendocrine function [21]. Locomotor responses to systemic administration of 10 mg/kg cocaine was reduced in ob/ob mice, while conditioned place preference was increased by 2.5 mg/kg of cocaine [22]. These results contributed new important information about METH effects on females, extending our group’s previous report [14].
Dopaminergic projections from the ventral tegmental area (VTA) to the nucleus accumbens (NAcc), and its control by the pedunculopontine nucleus [4], mediate psychostimulant stimulated locomotor activity [23]. Dopaminergic modulation is also required for feeding in ob/ob mice [7-9]. Exogenous leptin administration has been shown to reduce repetitive action potential discharge in VTA [24] and pedunculopontine neurons [25,26]. Therefore, in leptin-deficient ob/ob mice there may be inhibitory mechanisms that compensate for leptin-mediated inhibition. D2 receptors are required for locomotor sensitization [27]. Reduced striatal D2R binding described for ob/ob mice [10] might explain the lower locomotor activity reported here after 7 day METH administration. In conclusion, it may be suggested that lack of circulating leptin during development blunted the ob/ob response to METH administration.
Voltage-gated T-type calcium channels expressed in the somatosensory thalamocortical system have been involved in psychostimulant-induced locomotor activation [19,28]. Here, we have evaluated mRNA expression of a wide range of voltage-gated calcium channels (T and P/Q types), HCN2, and glutamate receptor subunits in the somatosensory thalamic complex after METH treatment in both WT and ob/ob mice. Higher mRNA expression levels for voltage-gated CaV2.1 and CaV3.3 calcium channels and NMDA-GluN1, and AMPA-GluA1 glutamate receptor subunits were observed in somatosensory thalamic neurons in ob/ob mice. METH-administration did not influence those changes. No significant changes were observed for CaV3.1 calcium channels. Increased mRNA levels were also observed for HCN2 channels in ob/ob mice. Interestinlgy, METH administration reduced HCN2 levels in the sensory thalamus of ob/ob mice. Further experiments are still needed in order to quantify protein levels changes of these channels/receptors mediated by METH.
These transcriptional alterations observed in thalamic nuclei from ob/ob mice might underlie previously described impaired sleep consolidation [29]. Indeed, CaV2.1 voltage-gated calcium channels are located in the dendrites of ventrobasal neurons and support gamma band oscillations that modulate membrane depolarization towards action potential threshold [30]. Thalamic reticular neurons express CaV3.2 and CaV3.3 T-type calcium channel subunits [31]. CaV3.1 subunits containing T-type calcium channels are expressed at both pre-synaptic thalamic reticular axon terminals and post-synaptic ventrobasal cell bodies [32], and are implicated in the transitions between waking and slow sleep sleep [33]. Moreover, HCN2 channels are involved in controlling thalamocortical activity, reducing the susceptibility to convulsant agents [34].
In addition, it was reported that somatosensory loss induces profound changes in motor adaptation and anticipation in humans [35]. Therefore, changes reported here on thalamic nuclei gene expression might underlie alterations in somatosensory processing that might in turn induce detrimental effects on locomotion. Further studies are needed on psychostimulant-mediated actions in this and other leptin resistance animal models to elucidate their effects on sensory and motivational brain networks.
Funtional implications for thalamocortical rhytmicity
Voltage-gated ionic channels are responsible for neuronal excitability, endowing them with autorhythmic membrane oscillatory capabilities [36]. Synaptic receptors mediate functional contacts between neurons, spreading oscillations throughout neuronal networks. In such networks, autorhythmic neurons may either act as oscillators/pacemakers or as resonators [36,37]. Changes described here for mRNA levels of voltage-gated ionic channels and synaptic receptors would affect neuronal oscillations in the CNS. Indeed, blocking or transgenic elimination of P/Q-type calcium channels have been described to prevent the generation of gamma band oscillations [30,37,38]. T-type and HCN channels are physically associated [39] to underlie low-frequency oscillatory activity at the thalamocortical level [33,38,39,40]. Indeed, T-type channels are key elements mediating the sensitivity to psychostimulants [19,28], consistent with the observed enhancement in low frequency oscillatory activity after METH [41,42] and changes in protein levels of T-type channels mediated by cocaine [43] administration. Further studies are needed to clarify METH-dependent alterations of voltage-gated ionic channels and glutamate receptors protein levels.
Dr. Bisagno has been authorized to study drug abuse substances in animal models by A.N.M.A.T. (National Board of Medicine Food and Medical Technology, Ministerio de Salud, Argentina).
This work was supported by grants from FONCYT-Agencia Nacional de Promoción Científica y Tecnológica (Argentina); Préstamo BID 1728 OC.AR. PICT-2012-1769, PICT 2012-0924, PICT 2014-2499, PICT 2015-2594, PICT 2016-1728 and UBACYT 2014-2017 #20120130101305BA. In addition, this work was supported by NIH award R01 NS020246, NIH award P20 GM103425 and P30 GM110702.
Authors report no financial conflict of interest, or otherwise, related directly or indirectly to this study.
2021 Copyright OAT. All rights reserv
- Zhang Y, Proenca R, Maffei M, Barone M, Leopold L, et al. (1994) Positional cloning of the mouse obese gene and its human homologue. Nature 372: 425-432. [Crossref]
- Ahima RS, Flier JS (2000) Leptin. Annu Rev Physiol 62: 413-437. [Crossref]
- Pelleymounter MA, Cullen MJ, Baker MB, Hecht R, Winters D, et al. (1995) Effects of the obese gene product on body weight regulation in ob/ob mice. Science 269: 540-543. [Crossref]
- Fulton S, Pissios P, Manchon RP, Stiles L, Frank L, et al. (2006) Leptin regulation of the mesoaccumbens dopamine pathway. Neuron 51: 811-822. [Crossref]
- Leshan RL, Opland DM, Louis GW, Leinninger GM, Patterson CM, et al. (2010) Ventral tegmental area leptin receptor neurons specifically project to and regulate cocaine- and amphetamine-regulated transcript neurons of the extended central amygdala. J Neurosci 30: 5713-5723. [Crossref]
- Udagawa J, Hatta T, Naora H, Otani H (2000) Expression of the long form of leptin receptor (Ob-Rb) mRNA in the brain of mouse embryos and newborn mice. Brain Res 868: 251-258. [Crossref]
- Leinninger GM, Jo YH, Leshan RL, Louis GW, Yang H, et al. (2009) Leptin acts via leptin receptor-expressing lateral hypothalamic neurons to modulate the mesolimbic dopamine system and suppress feeding. Cell Metab 10: 89-98. [Crossref]
- Szczypka MS, Rainey MA, Palmiter RD (2000) Dopamine is required for hyperphagia in Lep(ob/ob) mice. Nat Genet 25: 102-104. [Crossref]
- Palmiter RD (2007) Is dopamine a physiologically relevant mediator of feeding behavior? Trends Neurosci 30: 375-381. [Crossref]
- Pfaffly J, Michaelides M, Wang GJ, Pessin JE, Volkow ND, et al. (2010) Leptin increases striatal dopamine D2 receptor binding in leptin-deficient obese (ob/ob) mice. Synapse 64: 503-510. [Crossref]
- Cadet JL, Bisagno V (2013) The primacy of cognition in the manifestations of substance use disorders. Front Neurol 4: 189. [Crossref]
- Cadet JL, Bisagno V, Milroy CM (2014) Neuropathology of substance use disorders. Acta Neuropathol 127: 91-107. [Crossref]
- Raineri M, Peskin V, Goitia B, Taravini IR, Giorgeri S, et al. (2011) Attenuated methamphetamine induced neurotoxicity by modafinil administration in mice. Synapse 65: 1087-1098. [Crossref]
- Raineri M, Gonzalez B, Goitia B, Garcia-Rill E, Krasnova IN, et al. (2012) Modafinil abrogates methamphetamine-induced neuroinflammation and apoptotic effects in the mouse striatum. PLoS One 7: e46599. [Crossref]
- Gonzalez B, Raineri M, Cadet JL, García-Rill E, Urbano FJ, et al. (2014) Modafinil improves methamphetamine-induced object recognition deficits and restores prefrontal cortex ERK signaling in mice. Neuropharmacology 87:188-197. [Crossref]
- Bisagno V, González B, Urbano FJ (2016) Cognitive enhancers versus addictive psychostimulants: The good and bad side of dopamine on prefrontal cortical circuits. Pharmacol Res 109: 108-118. [Crossref]
- González B, Rivero-Echeto C, Muñiz JA, Cadet JL, García-Rill E, et al. (2016) Methamphetamine blunts Ca2+ currents and excitatory synaptic transmission through D1/5 receptor-mediated mechanisms in the mouse medial prefrontal cortex. Addiction Biol 21(3):589-602. [Crossref]
- Goitia B, Raineri M, González LE, Rozas JL, Garcia-Rill E, et al. (2013) Differential effects of methylphenidate and cocaine on GABA transmission in sensory thalamic nuclei. J Neurochem 124: 602-612. [Crossref]
- Bisagno V, Raineri M, Peskin V, Wikinski SI, Uchitel OD, et al. (2010) Effects of T-type calcium channel blockers on cocaine-induced hyperlocomotion and thalamocortical GABAergic abnormalities in mice. Psychopharmacology (Berlin) 212: 205-214. [Crossref]
- Finocchietto PV, Holod S, Barreyro F, Peralta JG, Alippe Y, et al. (2011) Defective leptin-AMP-dependent kinase pathway induces nitric oxide release and contributes to mitochondrial dysfunction and obesity in ob/ob mice. Antioxid Redox Signal 15: 2395-2406. [Crossref]
- Erickson JC, Hollopeter G, Palmiter RD (1996) Attenuation of the obesity syndrome of ob/ob mice by the loss of neuropeptide Y. Science 274: 1704-1707. [Crossref]
- Muelbl MJ, Nawarawong NN, Clancy PT, Nettesheim CE, Lim YW, et al. (2016) Responses to drugs of abuse and non-drug rewards in leptin deficient ob/ob mice. Psychopharmacology (Berl) 233: 2799-2811. [Crossref]
- Urbano FJ, Bisagno V, González B, Rivero-Echeto MC, Muñiz JA, et al. (2015) Pedunculopontine arousal system physiology-Effects of psychostimulant abuse. Sleep Sci 8: 162-168. [Crossref]
- Hommel JD, Trinko R, Sears RM, Georgescu D, Liu ZW, et al. (2006) Leptin receptor signaling in midbrain dopamine neurons regulates feeding. Neuron 51: 801-810. [Crossref]
- Beck P, Mahaffey S, Urbano FJ, Garcia-Rill E (2013) Role of G-proteins in the effects of leptin on pedunculopontine nucleus neurons. J Neurochem 126: 705-714. [Crossref]
- Beck P, Urbano FJ, Williams DK, Garcia-Rill E (2013) Effects of leptin on pedunculopontine nucleus (PPN) neurons. J Neural Transm (Vienna) 120: 1027-1038. [Crossref]
- Durieux PF, Schiffmann SN, de Kerchove d'Exaerde A (2012) Differential regulation of motor control and response to dopaminergic drugs by D1R and D2R neurons in distinct dorsal striatum subregions. EMBO J 31: 640-653. [Crossref]
- Gangarossa G, Laffray S, Bourinet E, Valjent E (2014) T-type calcium channel Cav3.2 deficient mice show elevated anxiety, impaired memory and reduced sensitivity to psychostimulants. Front Behav Neurosci 8: 92. [Crossref]
- Laposky AD, Shelton J, Bass J, Dugovic C, Perrino N, et al. (2006) Altered sleep regulation in leptin-deficient mice. Am J Physiol Regul Integr Comp Physiol 290: R894-903. [Crossref]
- Pedroarena C, Llinás R (1997) Dendritic calcium conductances generate high-frequency oscillation in thalamocortical neurons. Proc Natl Acad Sci USA 94: 724-728. [Crossref]
- Talley EM, Cribbs LL, Lee JH, Daud A, Perez-Reyes E, Bayliss DA (1999) Differential distribution of three members of a gene family encoding low voltage-activated (T-type) calcium channels. J Neurosci 19: 1895-1911. [Crossref]
- Kovacs K, Sik A, Ricketts C, Timofeev I (2010) Subcellular distribution of low-voltage activated T-type Ca2+ channel subunits (Ca(v)3.1 and Ca(v)3.3) in reticular thalamic neurons of the cat. J Neurosci Res 88: 448-460. [Crossref]
- Steriade M, Llinás RR (1988) The functional states of the thalamus and the associated neuronal interplay. Physiol Rev 68: 649-742. [Crossref]
- Ying SW, Kanda VA, Hu Z, Purtell K, King EC, et al. (2012) Targeted deletion of Kcne2 impairs HCN channel function in mouse thalamocortical circuits. PLoS One 7: e42756. [Crossref]
- Bunday KL, Bronstein AM (2009) Locomotor adaptation and aftereffects in patients with reduced somatosensory input due to peripheral neuropathy. J Neurophysiol 102: 3119-3128. [Crossref]
- Llinás RR (1988) The intrinsic electrophysiological properties of mammalian neurons: insights into central nervous system function. Science 242: 1654-1664. [Crossref]
- Garcia-Rill E, Kezunovic N, Hyde J, Simon C, Beck P, et al. (2013) Coherence and frequency in the reticular activating system (RAS). Sleep Med Rev 17: 227-238. [Crossref]
- Llinás RR, Choi S, Urbano FJ, Shin HS. (2007) Gamma-band deficiency and abnormal thalamocortical activity in P/Q-type channel mutant mice. Proc Natl Acad Sci USA 104: 17819-17824. [Crossref]
- Fan J, Gandini MA, Zhang FX, Chen L, Souza IA, et al. (2017) Down-regulation of T-type Cav3.2 channels by hyperpolarization-activated cyclic nucleotide-gated channel 1 (HCN1): Evidence of a signaling complex. Channels (Austin) 3:1-10. [Crossref]
- Huang Z, Lujan R, Kadurin I, Uebele VN, Renger JJ, et al. (2011) Presynaptic HCN1 channels regulate Cav3.2 activity and neurotransmission at select cortical synapses. Nat Neurosci 14: 478-486. [Crossref]
- Newton TF, Kalechstein AD, Hardy DJ, Cook IA, Nestor L, et al. (2004) Association between quantitative EEG and neurocognition in methamphetamine-dependent volunteers. Clin Neurophysiol 115:194-198. [Crossref]
- Janetsian SS, Linsenbardt DN, Lapish CC (2015) Memory impairment and alterations in prefrontal cortex gamma band activity following methamphetamine sensitization. Psychopharmacology (Berl) 232: 2083-2095. [Crossref]
- Urbano FJ, Bisagno V, Wikinski SI, Uchitel OD, Llinás RR (2009) Cocaine acute "binge" administration results in altered thalamocortical interactions in mice. Biol Psychiatry 66: 769-776. [Crossref]