(+)-Aeroplysinin-1 is a brominated alkaloid compound used by some sponges for defense due to its potent antibiotic effects. (+)-Aeroplysinin-1 has a wide spectrum of anti-tumoral action and behaves as a potent anti-angiogenic compound for bovine aortic endothelial cells. EGFR has been suggested as a molecular target of (+)-aeroplysinin-1 as an anti-tumor compound. We have previously shown that (+)-aeroplysinin-1 prevents the phosphorylation of Bad and this relates to the mitochondria-mediated induction of apoptosis by this compound. In the present study we have tested the potential of (+)-aeroplysinin-1 to inhibit a panel of protein kinase activities and the effects of (+)-aeroplysinin-1 treatment on Akt and Erk signalling pathways. Our results clearly show that (+)-aeroplysinin-1 does not inhibit EGFR, VEGFR, Akt, Erk and other 19 protein kinase activities in vitro but (+)-aeroplysinin-1 treatment causes an inhibition of both Akt and Erk phosphorylation in endothelial cells.
aeroplysinin-1, Akt signaling, angiogenesis, cancer Erk signaling
(+)-Aeroplysinin-1 is an optically active 1,2-dihydroarene-1,2-diol alkaloid produced by the marine sponge Verongia aerophoba, which use it as a chemical defense to protect them from invasion of bacterial pathogens after tissue injury [1-4]. The antibiotic action of aeroplysinin-1 makes it a potent antibacterial [1,4,5] and antiparasitic compound [6]. Furthermore, aeroplysinin-1 exhibits both cytostatic and cytotoxic effects on a wide range of tumor cells [7-11]. In particular, aeroplysinin-1 displays a potent anti-tumor effect on two human EGF-dependent breast cancer cell lines through its claimed inhibitory effect on EGF-receptor tyrosine kinase activity [8].
An alternative way to inhibit tumor progression is to block tumor angiogenesis [12,13]. We have previously characterized aeroplysinin-1 as a potent anti-angiogenic compound both in vitro and in vivo [14]. The anti-angiogenic effects of aeroplysinin-1 are connected with its induction of endothelial cell apoptosis by activation of the mitochondrial pathway [15].
To get further insight on the upstream signalling pathways leading to this pro-apoptotic effects of aeroplysinin-1 on endothelial cells, in the present work we have evaluated the potential of aeroplysinin-1 to inhibit protein kinase activities in vitro and to interfere with the phosphorylation of endothelial cell Akt and Erk, two key signalling molecules involved in the control of both cell proliferation and survival.
Our results suggest that the apoptosis-inducing mechanism of aeroplysinin-1 is endothelial cell-specific and dependent on disruption of Akt and Erk pathways.
Ethics statement
Primary cultures of HUVEC were obtained from umbilical cords donated at the Maternity of the University Clinical Hospital (Málaga) with the verbal informed consent of donors according to the procedure approved by the ethics committee. All personal data were maintained anonymous and the whole procedure remained anonymous to the authors of this article, who received the donated umbilical cords outside of the operating room. All the procedures were carried out following the rules provided by the bioethical committee of the University of Málaga. This study is part of a research project approved by the bioethical committee of the University of Málaga.
Chemicals and reagents
(+)-Aeroplysinin-1 was provided by Instituto Biomar (León, Spain) and was dissolved in DMSO and stored at -20°C. Anti-Akt, anti-phospho-Akt (Ser473), anti-p44/42 MAP kinase and anti-phospho-p44/42 MAPK (Thr202/Tyr204) rabbit polyclonal antibodies were purchased from Cell Signaling Technology. Rabbit anti-GAPDH antibody was purchased from Nordic BioSite. Goat anti-rabbit and anti-mouse secondary HRP-conjugated antibodies were from Pierce. Cell culture reagents were from Life Technologies. All other reagents were supplied by Sigma-Aldrich.
In vitro kinase inhibition assay
Kinase inhibition screening was a customized service provided by ProQinase GmbH (Freiburg, Germany).
Cell culture
Bovine aortal endothelial (BAE) cells were growth in Dulbecco’s modified Eagle’s medium (DMEM) containing 1 g/L glucose, 10% foetal calf serum, 2 mM glutamine, 50 U/mL penicillin, 50μg/mL streptomycin, 1.25 μg/mL amphotericin B. Human umbilical vein endothelial (HUVE) cells were isolated from umbilical cords and grown on gelatin-coated dishes in Medium 199 containing 10mM HEPES, 20% foetal calf serum, 2 mM glutamine, 100 μg/mL heparin and 30 μg/mL endothelial cell growth supplement (ECGS, Sigma-Aldrich, St Louis, MI, USA), 50 U/mL penicillin, 50 μg/mL streptomycin, 1.25 μg/mL amphotericin B. Passages three to seven were used for experiments. Human colon carcinoma cell line HCT-116 (a kind gift from Prof. B. Vogelstein, Johns Hopkins Kimmel Comprehensive Cancer Center), human fibrosarcoma HT-1080 and mouse embryonic fibroblast (MEF) were grown in DMEM containing 4.5 g/L glucose, 10% foetal calf serum, 2 mM glutamine, 50 U/mL penicillin, 50 μg/mL streptomycin, 1.25 μg/mL amphotericin B. All cell lines were maintained at 37°C and humidified 5% CO2 atmosphere.
Treatments
Aeroplysinin-1 was dissolved in DMSO and stored at -20°C. Cells were grown until 75% confluence. Afterwards, cells were starved overnight by serum deprivation and treated for 2 hours with aeroplysinin-1. Then, cells were stimulated for 10 min with medium containing 10% FBS and harvested for analysis.
Western-blot
Cells were lysed in Laemmli’s loading buffer 2X and boiled for 5 min at 95°C. Samples were separated by SDS-PAG electrophoresis and blotted onto nitrocellulose membranes using standard procedures. After blocking in TBS-T plus 5% non-fat dry milk, membranes were probed with primary antibodies overnight at 4°C. Then, the membranes were washed in TBS-T and probed with horseradish peroxidase (HRP)-conjugated secondary antibodies in blocking solution for 1 hour at room temperature. After washing, membranes were developed using the ECLTM system (Amersham Biosciences). For antibody re-probing, membranes were incubated in stripping solution (62.5 mM Tris-HCl pH 6.8, 2% SDS and 0.77% beta-mercaptoethanol) for 30 min at 50°C with shaking.
(+)-Aeroplysinin-1 does not inhibit kinase activities in vitro
The inhibitory effect on EGFR tyrosine kinase activity has been suggested as a direct molecular target for the anti-tumor effects of aeroplysinin-1 [8]. Since receptor tyrosine kinases and other protein kinases play key roles in the angiogenic process [16], we decided to carry out an in vitro kinase inhibition screening testing the effect of both 2 and 20 μM (+)-aeroplysinin-1 on the in vitro activity of the 25 protein kinases listed in Table 1. Even at the highest tested concentration (20 μM), (+)-aeroplysinin-1 was unable to produce a significant inhibitory effect on any of these protein kinase activities. In particular, EGFR and VEGFR2 activities remained unaltered.
Table 1. List of the 25 protein kinases included in the in vitro kinase inhibition screening testing the effect of both 2 and 20 µM (+)-aeroplysinin-1.
Protein kinases |
ABL1 |
AKT1 |
AKT2 |
ALK |
AMPK |
EGFR |
ERK1 |
ERK2 |
FAK |
FGFR1 |
FGFR2 |
FGFR3 |
FGFR4 |
IKK-beta |
KIT |
MET |
mTOR |
PDGFR-alpha |
PDGFR-beta |
TGFR1 |
TGFR2 |
TIE2 |
VEGFR1 |
VEGFR2 |
VEGFR3 |
(+)-Aeroplysinin-1 treatment inhibits Akt and Erk phosphorylation in endothelial cells
The control of survival pathways is directly involved in the regulation of endothelial apoptosis [16]. The effect of (+)-aeroplysinin-1 on cell survival pathways was analyzed in endothelial (BAE and HUVE), tumoral (HCT-116 and HT-1018) and non-transformed (MEF) cells. Cells were starved overnight by serum deprivation and treated with 10 and 20 µM (+)-aeroplysinin-1 for 2 hours or left untreated. Then, they were stimulated with serum, with exception of the negative control. As shown in Figure 1, phosphorylation of both Akt and Erk in BAE cells was significantly abrogated by 20 μM (+)-aeroplysinin-1, reaching levels similar to or below those of the negative controls. Similar results were obtained with HUVE cells in which level of Akt dephosphorylation exhibited a dose-dependent response. On the contrary, (+)-aeroplysinin-1 was unable to thwart Akt and Erk phosphorylation in the tumour cell lines HCT-116 and HT-1018. Moreover, treatment of MEF cells (that harbour intact Akt and Erk pathways) with (+)-aeroplysinin-1 had no effect on the phosphorylation status of Akt or Erk (Figure 1). The differential effect of (+)-aeroplysinin-1 on different cell types reflects the selectivity of this drug to target specific pathways functioning in endothelial cells, which maybe defective or absent in the other cell lines used in this study.
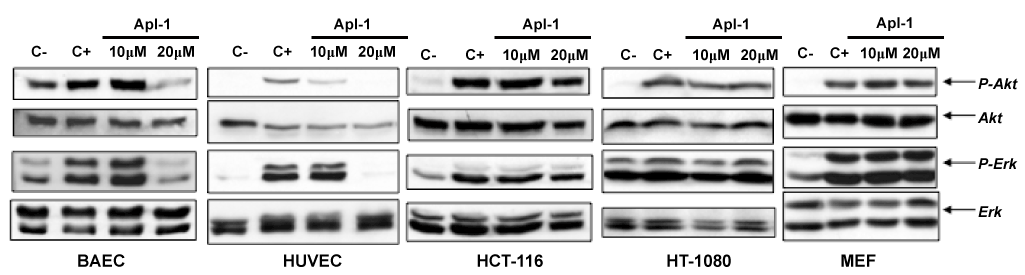
Figure 1. Effects of (+)-aeroplysinin-1 (Apl-1) on Akt and Erk phosphorylation in endothelial and non-endothelial cells. Cells starved overnight by serum deprivation were treated with Apl-1 for 2 hours and induced with serum during 10 minutes. Lysates prepared in Laemmli’s buffer were analyzed by Western blot for phosphorylation of Akt (P-Akt) and Erk (P-Erk). Membranes were reprobed for total Akt and Erk. Negative control (C-) corresponds to starved but not induced cells; positive control (C+) corresponds to starved and serum-induced cells. Identical results were obtained in two independent experiments.
The alkaloid compound aeroplysinin-1, synthesized by several marine sponges and secreted by them as a defensive weapon [1,4,5], exhibit cytostatic and cytotoxic effects on a wide range of both prokaryote and eukaryote cells [1,4-11,14,17]. A suggested molecular target for the anti-tumor effect of aeroplysinin-1 is the EGFR tyrosine kinase activity. Several reports provide evidence concerning the inhibitory effect of aeroplysinin-1 on EGF receptor [8], although this effect has been questioned by others [18]. A very recent docking study predicts that aeroplysinin-1 can interact with the EGFR catalytic kinase domain and suggests that similar interaction could be effective for inhibitory effects of aeroplysini-1 on other receptor tyrosine kinases such as VEGFR2, a key signalling molecule in angiongenesis [19]. Unfortunately, this does not seem to be the case in the real world according to the completely negative results obtained in our in vitro kinase inhibition screening testing the effect of both 2 and 20 μM (+)-aeroplysinin-1 on the in vitro activity of the 25 protein kinases listed in Table 1. In fact, this screening shows that (+)-aeroplysinin-1 is absolutely unable to inhibit any tested protein kinase activity, including those of EGFR and VEGFR2.
On the other hand, the anti-angiogenic effects described for aeroplysinin-1 include its potent inhibitory effect on endothelial cell proliferation (with an IC50 value of 2 µM for BAEC) and its induction of endothelial apoptosis by activation of the mitochondrial pathway. Our previously published results suggest that multiple targets ate involved in the activating effect of (+)-aeroplysinin-1 on endothelial cell apoptosis. They include Bad dephosphorylation and translocation to mitochondria, where its dimerization with Bcl-XL would promote the release of cytochrome c leading to caspase activation [15]. Among the network of pathways governing angiogenesis, the extracellular signal-related kinases-mitogen-activated protein kinases (ERK-MAPK) cascade and the PI3K/Akt pathway play a critical role, controlling the transduction of both proliferation and survival signals [16]. Interestingly, both signalling pathways are upstream of Bad phosphorylation (Figure 2). The capability of (+)-aeroplysinin-1 to inhibit the phosphorylation of Erk and Akt (as clearly shown in Figure 1) could account for its anti-angiogenic activity. However, the negative results obtained in our in vitro kinase inhibition screening show that (+)-aeroplysinin-1 is also unable to inhibit in vitro Akt and Erk kinase activities directly. The present work supports the hypothesis proposing (+)-aeroplysinin-1 as transduction pathways modulator. Interestingly, according to our results, aeroplysinin-1 seems to have a specific effect on endothelial cells, since no effect is detected in the transduction pathways of tumour cell lines HCT-116 and HT-1080 (Figure 1), where those signalling pathways are deregulated [20,21]. Similarly occurs in MEF cells, although they have intact signalling pathways. This feature points to aeroplysinin-1 as a putative inhibitor of a specific unidentified mediator downstream of some endothelial tyrosine kinase receptor involved in the activation of both PI3K/Akt and MAPK pathways. Thus, the blockage of this common mediator would result in the inhibition of both routes. This hypothesis would explain why aeroplysinin-1 does selectively inhibit Erk and Akt phosphorylation in endothelial cells. However, further studies are needed to test this hypothesis. Figure 2 summarizes the current state of knowledge and suggest the connection of the results of the present work with those previously published by our group showing tha activating effect of (+)-aeroplysinin-1 on endothelial cell apoptosis through inhibition of Bad phosphorylation, stimulation of cytochrome c release and activation of caspases [15].
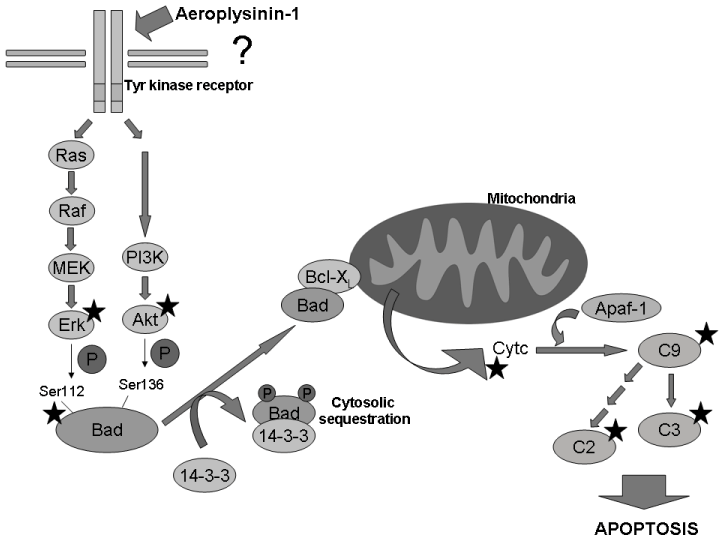
Figure 2. Apoptosis induction mechanism proposed for Apl-1. Putative inhibition of an unidentified mediator downstream of some tyrosine-kinase receptor specific of endothelial cells leads to the inhibition of the phosphorylation of both Erk and Akt. In these circumstances, Bad would be dephosphorylated and translocated from cytosol to mitochondria, where it would dimeryze with Bcl-XL, thus promoting the release of cytochrome c, activating caspase 9 and subsequent apoptosis (results for Apl-1 in endothelial cells obtained in this work and in our previous article [15] are indicated with a star).
Authors are indebted to Auxiliadora López Jiménez for her excellent technical assistance. Thanks are due to Instituto Biomar S.A. (León, Spain) for providing us the compound.
This work was supported by grants BIO2014-56092-R (MINECO and FEDER) and P12-CTS-1507 (Andalusian Government and FEDER). The “CIBER de EnfermedadesRaras” is an initiative from the ISCIII (Spain). The funders had no role in the study design, data collection and analysis, decision to publish or preparation of the manuscript.
Conceived and designed the experiments: Ana R. Quesada, Miguel Ángel Medina.
Performed the experiments: Beatriz Martínez Poveda.
Analyzed the data: Beatriz Martínez Poveda, Ana R. Quesada, Miguel Ángel Medina.
Wrote the paper: Miguel Ángel Medina.
- Fattorusso E, Minale L, Sodano G (1972) Aeroplysinin-1, an antibacterial bromo-compound from the sponge Verongia aerophoba. J Chem Soc Perkin 1 1: 16-18. [Crossref]
- Debitus C, Guella G2021 Copyright OAT. All rights reserv et al. (1998) Quinolones from a bacterium and tyrosine metabolites from its host sponge, Suberea creba from the Coral Sea. J Mar Biotechnol 6: 136-141. [Crossref]
- Thoms C, Wolff M, Padmakumar K, Ebel R, Proksch P (2004) Chemical defense of Mediterranean sponges Aplysina cavernicola and Aplysina aerophoba. Z Naturforsch C 59: 113-122. [Crossref]
- Thoms C, Ebel R, Proksch P (2006) Activated chemical defense in aplysina sponges revisited. J Chem Ecol 32: 97-123. [Crossref]
- Teeyapant R, Woerdenbag HJ, Kreis P, Hacker J, Wray V, et al. (1993) Antibiotic and cytotoxic activity of brominated compounds from the marine sponge Verongia aerophoba. Z Naturforsch C 48: 939-945. [Crossref]
- Galeano E, Thomas OP, Robledo S, Munoz D, Martinez A (2011) Antiparasitic bromotyrosine derivatives from the marine sponge Verongula rigida. Mar Drugs 9: 1902-1913. [Crossref]
- Kreuter MH, Bernd A, Holzmann H, Müller-Klieser W, Maidhof A, et al. (1989) Cytostatic activity of aeroplysinin-1 against lymphoma and epithelioma cells. Z Naturforsch C 44: 680-688. [Crossref]
- Kreuter MH, Leake RE, Rinaldi F, Muller-Klieser W, Maidhof A, et al. (1990) Inhibition of intrinsic protein tyrosine kinase activity of EGF-receptor kinase complex from human breast cancer cells by the marine sponge metabolite (+)-aeroplysinin-1. Comp Biochem Physiol B 97: 151-158. [Crossref]
- Kreuter MH, Robitzki A, Chang S, Steffen R, Michaelis M, et al. (1992) Production of the cytostatic agent aeroplysinin by the sponge Verongia aerophoba in in vitro culture. Comp Biochem Physiol C 101: 183-187. [Crossref]
- Koulman A, Proksch P, Ebel R, Beekman AC, van Uden W, et al. (1996) Cytotoxicity and mode of action of aeroplysinin-1 and a related dienonefrom the sponge Aplysina aerophoba. J Nat Prod 59: 591-594. [Crossref]
- Stuhldreier F, Kassel S, Schumacher L, Wesselborg S, Proksch P, et al. (2015) Pleiotropic effects of spongean alkaloids on mechanisms of cell death, cell cycle progression and DNA damage response (DDR) of acute myeloid leukemia (AML) cells. Cancer Lett 361: 39-48. [Crossref]
- Carmeliet P (2005) Angiogenesis in life, disease and medicine. Nature 438: 932-936. [Crossref]
- Folkman J (2007) Angiogenesis: an organizing principle for drug discovery? Nat Rev Drug Discov 6: 273-286. [Crossref]
- Rodríguez-Nieto S, González-Iriarte M, Carmona R, Muñoz-Chápuli R, Medina MA, et al. (2002) Antiangiogenic activity of aeroplysinin-1, a brominated compound isolated from a marine sponge. FASEB J 16: 261-263. [Crossref]
- Martínez-Poveda B, Rodríguez-Nieto S, García-Caballero M, Medina MÁ, Quesada AR (2012) The antiangiogenic compound aeroplysinin-1 induces apoptosis in endothelial cells by activating the mitochondrial pathway. Mar Drugs 10: 2033-2046. [Crossref]
- Muñoz-Chápuli R, Quesada AR, Medina MÁ (2004) Angiogenesis and signal transduction in endothelial cells. Cell Mol Life Sci 61: 2224-2243. [Crossref]
- Martinez-Poveda B, Garcia-Vilas JA, Cardenas C, Melgarejo E, Quesada AR, et al. (2013) The brominated compound aeroplysinin-1 inhibits proliferation and the expression of key pro- inflammatory molecules in human endothelial and monocyte cells. PLoS One 8: e55203. [Crossref]
- Hinterding K, Knebel A, Herrlich P, Waldmann H (1998) Synthesis and biological evaluation of aeroplysinin analogues: a new class of receptor tyrosine kinase inhibitors. Bioorg Med Chem 6: 1153-1162. [Crossref]
- Garcia-Godoy MJ, Lopez-Camacho E, Garcia-Nieto J, Aldana-Montes AJ (2015) Solving molecular docking problems with multi-objective metaheuristics. Molecules 20: 10154-10183. [Crossref]
- Brown R, Marshall CJ, Pennie SG, Hall A (1984) Mechanism of activation of an N-ras gene in the human fibrosarcoma cell line HT1080. EMBO J 3: 1321-1326. [Crossref]
- Shirasawa S, Furuse M, Yokoyama N, Sasazuki T (1993) Altered growth of human colon cancer cell lines disrupted at activated Ki-ras. Science 260: 85-88. [Crossref]